

Ekologiya
.pdfрізних частинах дерев і характеризується деякими відмінностями відшукування комах серед гілок і листя.
Важливим засобом "заспокоєння" лімітуючої конкуренції, яка врештірешт переходить у співіснування, є кормова спеціалізація, яка часто базується на розмірах жертви: більший хижак поїдає більшу жертву, менший – меншу.
Англійський еколог Д.Лек (1971) описав співіснування п'яти видів синиць у широколистяних лісах поблизу Оксфорда і дійшов висновку, що протягом більшої частини року вони відокремлені із-за відокремленості їхніх кормових ділянок, відміни в розмірах комах і міцності насіння, яким вони живляться. Екологічна відокремленість пов'язана з відмінністю в масі синиць, розмірах і формі дзьоба. Незважаючи на подібність синиць, кожний вид по-різному використовує свої кормові ресурси. Велика синиця годується переважно на землі, поїдаючи комах довжиною понад 6 мм, лісові горіхи, жолуді, насіння пшениці, букові горішки. Болотяна гаїчка тримається вище, ніж велика синиця, але нижче голубої синиці, годуючись у чагарнику, в нижніх ярусах дерев і на траві комахами розміром 3-4 мм, насінням лопуха, бруслини, жимолості та кислиці. Вертка невелика голуба синиця годується переважно в кронах дубів, оскільки мала маса та спритність дають їй змогу втримуватися на дрібних гілочках і листках. До її раціону входять комахи, розміри яких звичайно не перевищують 2 мм. Вона дістає їх з-під кори. Як правило, голуба синиця не живиться насінням (за винятком березового). Московка, на відміну від лазорівки, тримається частіше всього великих гілок дуба або ялини, які відходять від стовбура. Живиться переважно комахами довжиною менше 2 мм. І нарешті, буроголова гаїчка, яка дуже подібна до болотяної гаїчки, годується на березі, бузині і трав'яному покриві; на відміну від болотяної гаїчки практично не зустрічається на дубах, поїдає дуже мале насіння.
М.Бігон, Дж.Харпер і К.Таунсенд (1991) подають три можливі варіанти пояснення такого співіснування видів.
Перше базується на так званій "поточній конкуренції" (сиrепt сотреtitiоп). Наприклад, синиці є конкуруючими видами, їх співіснування є наслідком роздільності екологічних ніш. Однак у випадку відсутності конкурента вони можуть розширити свої ніші, тобто освоювати фундаментальні ніші.
Друге – це еволюційно зумовлене уникання конкуренції, яке Коннел (Соппеll, 1980) назвав "привидом конкурентного минулого". П'ять згаданих вище видів синиць, домівкою яких стали ліси поблизу Оксфорда, давно вже "притерлися" між собою, і їх конкуренція залишилася в далекому еволюційному минулому, їхні фундаментальні екологічні ніші вже давно перекриваються.
83
Третє пояснення також можна обґрунтувати тією ж ситуацією з синицями. В ході своєї еволюції ці види синиць по-різному і незалежно один від одного реагували на природний відбір, оскільки це різні види, які мають різні ознаки. Однак вони не конкурують у даний момент і ніколи не конкурували в минулому, тому що просто виявились різними.
Немає сумніву в тому, що ці три пояснення, взяті разом чи зокрема, не можуть однозначно витлумачити якийсь із наведених прикладів співіснування видів. Еколог має провести чимало аналітичних розрахунків, щоб встановити, яке із трьох пояснень може бути ймовірним для конкретної ситуації.
Еволюційна дивергенція. При співіснуванні подібних видів міжвидова конкуренція сприяє розвиткові екологічної дивергенції (роз'єднання), яка зменшує можливості перекриття під час використання ресурсів. При неповному перекритті ареалів двох видів міжвидова конкуренція впливає на еволюцію у зоні перекриття, але не в інших місцях.
Про еволюційну дивергенцію можна говорити в тих випадках, коли вид у присутності конкурента виявляє зовнішні ознаки або особливості поведінки, які відрізняються від таких в умовах відсутності конкуренції. Це явище, яке називають зміщенням ознак (наприклад, різкі розміри дзьоба), ще раз свідчить, що конкуренція може зумовити еволюційну дивергенцію.
Конкурентне виключання й еволюційна дивергенція зменшують міжвидову конкуренцію, забезпечуючи тим самим ефективне використання ресурсів і підтримку загального потоку енергії та поживних речовин в екосистемі на найвищому рівні, який лише можливий при даному складі угруповання.
2.5.3 Хижацтво
Якщо конкуренція проявляється у взаємному впливі конкуруючих видів, то хижацтво – це однобічний процес, який характеризує стосунки між хижаком і жертвою.
Хижаки – це тварини або рослини, які полюють і поїдають жертву.
Як правило, вони мають широке коло живлення, можуть мобільно переключатися з одної здобичі на іншу – доступнішу та чисельнішу. З екологічної точки зору стосунки хижак – жертва є сприятливі для одного виду і несприятливі для іншого. Водночас обидва види формують такий спосіб життя і таке чисельне співвідношення, які врешті-решт забезпечать їм нормальне співіснування (популяції вовка і оленя).
Особливістю хижацтва є те, що в цих стосунках один вид витрачає багато кмітливості і енергії, щоби схопити і з'їсти жертву, а інший вид – щоби утекти. Перший і другий у процесі тривалої еволюції екологічно адаптувалися; хижак розвинув такі якості, як гострота органів чуття,
84
блискавична реакція і швидкий біг та інше, жертва, в свою чергу – захисне (камуфляжне) забарвлення, панцирі, шипи, голки, отруйні викиди тощо. Співвідношення особин популяцій хижака і жертви, як правило, є таким, що забезпечує безмежно тривале співіснування видів, а отже, і біологічну регуляцію популяцій.
Типовими хижаками, наприклад серед ссавців, є котячі, вовк, лисиця, горностай, куниця, тюлені, моржі; поміж птахів – орел, яструб, сокіл; поміж риб – щука, окунь, форель. Є хижі рептилії (крокодил, алігатор), комахи і навіть гриби.
Наслідки впливу хижаків на стан популяції жертви можуть проявлятися не відразу, а через деякий час. Тому виникає багато питань щодо способів впливу хижаків на їх жертви і наслідків взаємостосунків для угруповання в цілому. Наприклад, доки хижаки стабілізують популяцію жертви або, навпаки, зумовлюють коливання її чисельності.
Вплив хижаків на популяцію жертви. Модель Лотки – Вольтерра.
Модель для хижака і жерти була створена в 20-х роках Лоткою і Вольтерра. Складається вона з двох компонентів: С – чисельність популяції консумента (хижака) і N – чисельність або біомаса популяції жертви чи рослин.
Спочатку розглянемо варіант, коли за відсутності хижака популяція жертви буде зростати за експоненціальною кривою:
dN |
rN . |
(2.16) |
|
dt |
|||
|
|
Зрозуміло, що хижаки знищують жертв зі швидкістю, яка визначається частотою зустрічей особин хижака і жертви, а частота зустрічей зростає в міру збільшення чисельності хижака (С) і жертви (N). Проте точна кількість зустрінутих і з'їдених жертв буде залежати від ефективності, з якою хижак знаходить і ловить жертву, тобто від а1 – "ефективності пошуку" або "частоти нападів". Отже, швидкість поїдання жертви буде α1∙ C·N або ж
dN |
rN 1 C N . |
(2.17) |
|
dt |
|||
|
|
Настає певний момент, коли через відсутність їжі окремі особини хижаків втрачають масу, голодують і гинуть. Припустимо, що у нашій моделі чисельність популяції хижака внаслідок голодування буде зменшуватися експоненціально:
dN |
|
C N , |
(2.18) |
|
|||
|
|||
dt rN a |
|||
|
85 |
|
|

dC |
q C , |
(2.19) |
|
dt |
|||
|
|
де q – смертність.
Загибель компенсується народженням нових особин зі швидкістю, яка, як береться до уваги в даній моделі, залежить лише від двох параметрів: 1) швидкості споживання їжі, ; 2) ефективності f з якою ця їжа переходить у потомство хижака. Отже, народжуваність хижака дорівнює f a C N , і в цілому становить:
dC |
|
C N q C . |
(2.20) |
|
|||
dt f a |
|||
Рівняння 2.18 і 2.20 становлять модель Лотки -Вольтерра. Властивості цієї моделі можна дослідити, побудувавши ізокліни –
лінії, які відповідають постійній чисельності популяції. За допомогою таких ліній визначають поведінку взаємодіючих популяцій хижак-жертва. У випадку із популяцією жертви (рівняння 2.18):
dN |
|
|
|
r |
||
|
|
|
|
|
|
|
dt 0 , |
|
або C a . |
||||
r N a |
C N |
|||||
Оскільки r і а' константи, ізокліною для жертви буде лінія, для якої величина С є постійною (рис.2.7, а).
Для хижаків ситуація буде аналогічною (2.20):
dC |
0 , |
f a C N q C |
або |
N |
q |
, |
(2.21) |
|
dt |
f a |
|||||||
|
|
|
|
|
|
тобто ізокліною для хижака буде лінія, вздовж якої N постійна (рис. 2.7,б). Ці рівняння надзвичайно прості для виявлення реальної взаємодії хижак-жертва, оскільки в основному спрямовані на виявлення циклічних коливань чисельності обох популяцій і не беруть до уваги можливе
існування стабільної популяції.
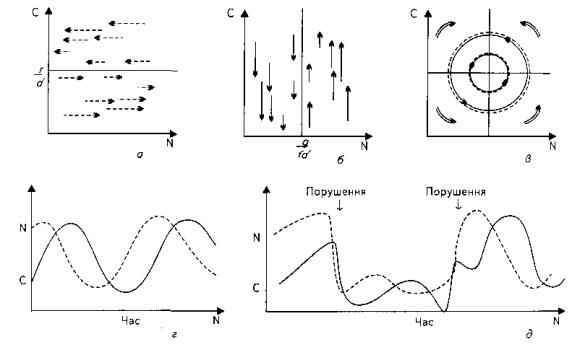
Якщо помістити обидві ізокліни на одному рисунку (рис.2.7,в), то наглядно побачимо взаємодію популяцій, коливання їх чисельності, які часто мають вигляд тісно пов'язаних між собою циклів. У кожному циклі, як тільки збільшується популяція жертви, починає рости і популяція хижака (рис.2.7,г). Вона перевищує чисельність жертви, а для підтримування стабільності популяції "переїдає" свій кормовий ресурс, понижуючи розмір популяції жертви.
Модель рівноваги системи хижак-жертва. Подібно як показники народжуваності та смертності розташовані в основі моделі росту
86
чисельності популяції, так показники поповнення (за рахунок розмноження чи імміграції) та виїдання жертви розташовані в основі
моделі рівноваги системи хижак-жертва. Якщо поповнення перевищує виїдання, то показник жертви збільшується, і навпаки, якщо виїдання перевищує поповнення, то він скорочується. Точки перетину кривих поповнення і виїдання є одночасно точками стійкої рівноваги для популяції жертви (рис. 2.8)
Рис..2.7. – Модель Лотки-Вольтерра для системи хижак-жертва:
а-ізокліна для популяції жертви. При низькій щільності хижака (С) чисельність жертви (N) зростає, а при більш високій зменшується; б- ізокліна популяції хижака. При високій щільності жертви чисельність популяції хижака зростає, а при низький зменшується (в). При об’єднанні ізоклін у системі хижак-жертва виникають необмежені взаємопов’язані коливання чисельності , які подібні до тих, які показані на рис.г. Однак для цих коливань (д) характерна нейтральна стабільність: при відсутності порушень вони тривають необмежено довго, але після кожного порушення, що призводить до нового рівняння чисельності, починається нова серія нейтрально стабільних циклів.
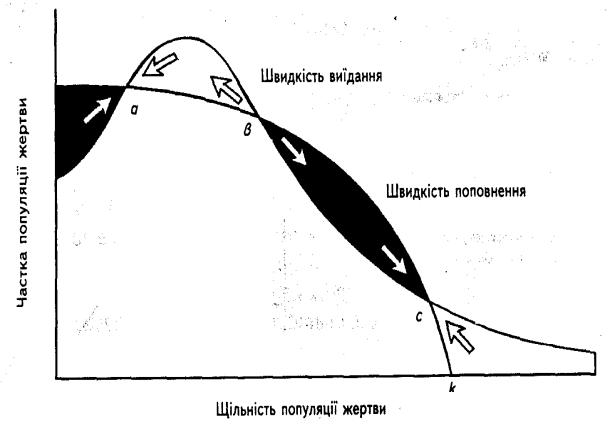
Таких точок три: верхня та нижня є точками стійкої рівноваги, поблизу яких відбувається регуляція чисельності популяції; середня – це точка нестійкої проміжної рівноваги, через яку популяція проходить на шляху від однієї точки стійкої рівноваги до іншої.
Точка рівноваги відповідає ситуації, за якої популяція жертви регулюється головним чином кормовими й іншими ресурсами середовища, а вплив хижаків має другорядний характер. У точці а хижаки
87
доводять чисельність популяції жертви до рівня, який менший від ємності середовища.
Рис.2.8 – Швидкість видання популяції жертви і її поповнення в гіпотетичній системі хижак-жертва. Чорним показано чисте зростання
чисельності жертви, а білим – її скорочення. Коли видання перевищує поповнення , популяція жертви скорочується, і навпаки (показано стрілками). Точки а і с відповідають точкам стійкої рівноваги для популяції жертви; точка а відображає регуляцію чисельності популяції хижаками; точка с – регуляцію харчовими та іншими ресурсами; k – координатна вісь.
До речі, вплив хижаків на популяцію жертви у точці а досить нестійкий. Якщо сильні морози або іззовні занесена хвороба зменшать популяцію хижака і втримають її на низькому рівні протягом тривалого часу, за яким популяція жертви встигне перейти за точку b, то зростання її чисельності буде тривати незалежно від того, відбудеться чи не відбудеться відновлення популяції хижака. Ця ситуація досить часто трапляється в сільському господарстві, де шкідник, чисельність якого звичайно втримується на безпечному для посівів рівні хижаками і паразитами, дає раптово загрозливий спалах. Після такого спалаху хижаки (наприклад, птахи) вже не в змозі регулювати популяцію шкідника доти, доки яка-небудь подія в середовищі (хоча б внесення пестицидів) не
88
знищить чисельність цієї популяції, тобто не поверне її в сферу впливу хижака.
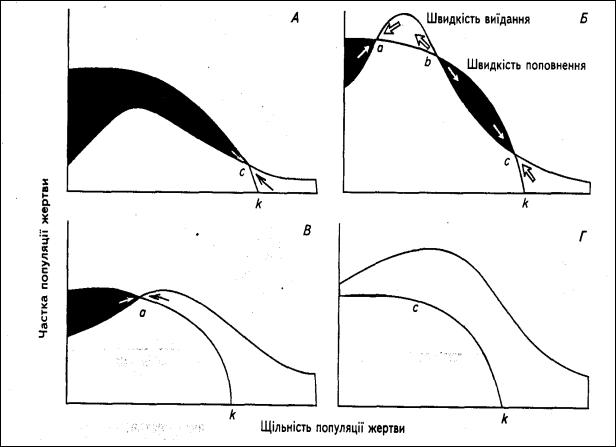
Користуючись діаграмою швидкостей виїдання і поповнення жертви (рис.2.8), можна прогнозувати наслідки різних рівнів впливу хижака на чисельність популяції жертви (рис.2.9). Неефективні хижаки не можуть регулювати популяції жертви при низькій її щільності: вони несуттєво зменшують чисельність жертви, але її популяція залишатиметься поблизу рівня рівноваги, який встановлюється наявними в середовищі ресурсами (рис.2.9, А, точка с). Зростання ефективності хижаків при низькій щільності жертви може привести до регулювання з боку хижака у точці
а (рис.2.9, Б).
Рис. 2.9 – Криві видання і поповнення популяції жертви при різних
інтенсивностях хижацтва. Чорним вказано чисте зростання чисельності жертви, а білим – її скорочення.
Якщо функціональні та чисельні реакції достатні для підтримки щільностей хижаків або ж якщо чисельність жертви порівняно з рівнем хижаків обмежена низькою ємністю середовища, то хижаки можуть ефективно регулювати популяцію жертви в будь-яких обставинах, і тоді
89
точка рівноваги зникає (рис.2.9,В). Розглянемо і таку ситуацію, коли внаслідок інтенсивного хижацтва популяція жертви виїдається до повного зникнення (рис.2.9,Г) (точка рівноваги відсутня). Подібну ситуацію можна створити в лабораторних умовах, оскільки в природному середовищі це можливо лише тоді, коли хижак має якийсь альтернативний корм. У біологічних методах боротьби зі шкідниками, на думку екологів, ефективність використання хижаків і паразитів даного шкідника можна підвищити, постачаючи їх якою-небудь альтернативною жертвою.
Таксономічна і функціональна класифікації хижаків. Вивчаючи різні варіанти взаємодії в системі хижак-жертва, бачимо, що одні хижаки живляться тваринами, інші – рослинами, а треті – відразу обома типами організмів. На цій конкуренції базується так звана таксономічна класифікація. Згідно з нею першу групу хижаків називають м'ясоїдними, другу – рослиноїдними, а третю – всеїдними.
Функціональна класифікація включає чотири різновиди хижаків: а)хижаки; 6) хижаки з пасовищним типом живлення; в) паразитоїди; г)паразити (мікро - і макропаразити).
Справжні хижаки вбивають свою жертву і з'їдають її повністю або ж частково. Це не тільки такі відомі хижаки, як тигри, вовки, орли, але й гризуни, які живляться насінням, а також мурашки, кити, що фільтрують планктон.
Хижаки з пасовищним типом живлення упродовж свого життя, як і перша група, з'їдають багато жертв (повністю або частково). Як правило, їх вплив на жертви є шкідливим. Сюди належать крупні травоїдні хребетні, такі як бізони, буйволи, олені, антилопи, вівці, а також свійська велика рогата худоба. Однак, відповідно до визначення, до хижаків з пасовищним типом живлення належать і мухи, які ссуть у них кров.
2.5.4 Паразитизм
Враховуючи особливості функціональної дії паразитів, розглянемо їх окремо. Паразит – організм, який живе на поверхні або в органах і тканинах інших організмів (живителів), живиться за їх рахунок, одержуючи від них готові речовини, та завдає їм певної шкоди. Розрізняють ендопаразитів, що селяться всередині живителя, і ектопаразитів, що існують на поверхні організму живителя. Виділяють також облігатні та факультативні форми паразитизму.
Паразити, подібно до хижаків з пасовищним типом живлення, поїдають лише частини своєї жертви (господаря). З того часу, – підкреслює Р.Уіттекер, – як паразит став постійно залежати від свого господаря як джерела живлення, йому стало невигідно убивати його. На відміну від справжніх хижаків і хижаків із пасовищним типом живлення, які вбивають велику кількість жертв, паразит тісно пов'язаний зі своєю
90
жертвою-господарем. Типовим прикладом паразита є туберкульозна паличка, яка "роз'їдає" легені ослабленого людського організму. Стосовно дерева, наприклад берези, паразитом є гриб - березовий трутовик, який своїми гіфами висмоктує з неї поживні речовини.
Паразитоїдами називають групу комах, головним чином перетинчастокрилих (Нутепоptеrа),а також багатьох двокрилих (Diptera) в яких дорослі самки і їхні личинки вирізняються подібністю поведінки.
Комахи-паразитоїди відкладають яйця в тіло іншої комахи або ж на його поверхню, тоді як комахи-господарі, як правило, ще не досягли дорослого віку. На їх тілі (або на їх поверхні, або поряд) із вилупленого яйця з'являється личинка, яка починає інтенсивно поїдати свого молодого господаря. Таким чином, господар гине, не досягши зрілого стану – в стадії лялечки або й раніше (яскравим прикладом такої поведінки є оса Chrisis ignata, яка паразитує на бджолах).
Паразитоїди тісно пов'язані з певним організмом господаря (подібно до паразитів), вони не призводять до швидкої загибелі господаря (подібно до паразитів і хижаків із пасовищним типом живлення), однак з часом господар неминуче загине (цим паразитоїди нагадують справжніх хижаків). Виділення паразитоїдів у окрему групу є цілком виправданим: за деякими даними до них належить близько 25% усіх видів, які живуть сьогодні на нашій планеті. Оскільки один вид комах має як мінімум одного паразитоїда, а останній – свого власного паразитоїда, то це свідчить про величезну кількість паразитоїдів.
Екологи добре вивчили природу цієї групи комах, використовують їх для боротьби зі шкідниками. Ю.Одум (1986) наводить приклад використання одного з видів хижого кліща, який ефективно контролює чисельність рослиноїдного кліща на посадках суниць у Каліфорнії. Водночас автор звертає увагу на неефективність використання неспеціалізованих хижаків, а часто й на непередбачене лихо, яке можуть спричинити некваліфіковані заходи біологічного захисту. Наприклад, мангусти, яких завезли на острови Карибського басейну для боротьби зі щурами на плантаціях цукрової тростини, почали масово поїдати птахів, які гніздилися на землі. Чисельність цих птахів зменшилась більше, ніж чисельність щурів.
2.5.5 Алелопатія
Алелопатія (або антибіоз) – один із прикладів аменсалізму. Це явище взаємного антагонізму серед рослин, яке виявляється у виділенні хімічних речовин, отруйних для іншого виду і не шкідливих для виду, що їх виділяє. Хімічні виділення є продуктами метаболізму – обміну речовин: ефірні олії, фітонциди, глюкозиди, які часто називають колінами.
91
Найчастіше алелопатія проявляється у конкурентному витісненні одного виду іншим. Ще в давнину помітили, що під горіхом, як правило, не росте трав'яна рослинність. Подібну властивість проявляє дуб звичайний
(Quercus robur) і особливо дуб північний (Quercus borealis). Пирій та інші бур'яни витісняють культурні рослини. Листя каштанів, яке розкладається, виділяє токсичні речовини, здатні пригнічувати ріст сіянців багатьох рослин.
Алелопатія суттєво впливає на швидкість рослинних сукцесій і на видовий склад стабільних угруповань. Хімічні виділення можуть впливати двояко: в одних випадках вони зменшують видову різноманітність, в інших (за рахунок адаптивних пристосувань, які складаються в умовах диференціації ніш) підтримують високу видову різноманітність.
Не завжди рослина виділяє токсини за допомогою коріння. Вважають малоймовірним, наприклад, що шавлія витісняє із угруповання інший вид саме цим способом. Дослідники спостерігали, що довкола групи рослин шавлії утворюється гола ділянка, яка відокремлює ці рослини від сусідніх ділянок, вкритих травою. Коріння, як виявилося, доходить лише до оголеної ділянки і його не можна вважати агентом негативного впливу на цю територію. Виявляється, що листя шавлії продукує легкі терпени (клас органічних сполук, до яких належить камфора), які, на думку Р.Ріклефса, впливають на рослини, що ростуть поблизу, безпосередньо через атмосферу. Сильні дощі виводять ці токсини із атмосфери в ґрунт, знижуючи антибіотичну дію шавлії на інші види. Цим зумовлюється відсутність безпосередньої хімічної конкуренції в регіонах з вологим кліматом. Водночас терпени, які виробляє шавлія і які подавляють ріст інших рослин, у процесі еволюції вироблялися не як засіб конкуренції, а як аттракторний (приваблюючий) засіб для його опилювачів, зокрема бджіл.
Алелопатія відома не лише в світі вищих рослин, але й серед мікроорганізмів. Наприклад, гриб мікроміцет Pinicilium пригнічує розвиток багатьох бактерій своїми хімічними інгібіторними виділеннями.
2.5.6 Позитивна взаємодія: коменсалізм, протокооперація, мутуалізм
Життя організмів характеризується не лише боротьбою, але й і багатьма формами взаємовигідного існування, яке склалося в процесі еволюції. Це, як зазначено вище, коменсалізм, протокооперація і мутуалізм.
Позитивні взаємодії популяцій розглянемо в тій послідовності, в якій вони формувалися в процесі еволюції: коменсалізм (перевагу чи користь має одна популяція), протокооперація (користь одержують обидві
92