

Ekologiya
.pdfрозселяється, і з того, що "осіло", виростають рослини, які дають насіння обох типів.
Диморфізм типу "волоцюга-домувальник" надзвичайно характерний також для тлі (у цих комах є як крилате, так і безкриле потомство). Крилата і безкрила форми генетично тотожні, тому що диморфізм виникає на тій фазі популяційного циклу, коли спостерігається партєногенетичне розмноження. Вид потомства, яке дасть материнська особина (крилате, безкриле або ж те і інше), залежить, мабуть, від щільності популяції і якості корму.
Досить часто спостерігаємо послідовний поліморфізм листя в кроні дерева. В деяких рослин у різні сезони утворюються зовсім різні за будовою листки, які опадають зі зміною сезону. Наприклад, у період року, коли у грунті достатньо води, деякі чагарники пустель в Ізраїлі (зокрема, Теисrіит рolіит) утворюють сильно розсічені листки з тонкою кутикулою. Коли ж настає посушливий сезон, ці листки змінюють інші – більш дрібні, шкірясті, і нерозсічені, які, в свою чергу, опадаючи, часто-густо залишають після себе лише зелені колючки та шпичаки. Таким чиом, тут у листяному покриві спостерігаємо послідовний (тобто такий, який реалізується не одночасно, а за ходом вегетаційного сезону) поліморфізм; листки одного типу змінюють листки іншого типу – з меншими фотосинтетичною активністю, водопроникністю.
2.4.4 Розселення
Живі істоти, в які б вони не входили "популяційні об'єднання", мають здатність до переміщення з одного місця на інше. Це роблять не лише швидкокрилі птахи, але й, здавалось би, нерухомі молюски і навіть "приякорені" до ґрунтового субстрату рослини, розсилаючи у всі кінці повітрям, водою, за допомогою тварин і, що сьогодні надзвичайно поширено, за участю людини своє насіння.
Так відбувається процес розселення-переселення видів за межі свого ареалу, що приводить до заселення нових ділянок, тобто натуралізації, яка виявляється в здатності рослин і тварин приживатись і давати потомство в нових для них природних біоценозах.
Під розселенням найчастіше розуміють віддалення одних особин від інших шляхом активного або пасивного переміщення, виділяючи декілька його видів: а) віддалення насіння рослин або личинок морських зірок одне від одного і від своїх родичів; б) перебігання полівок з однієї ділянки лугу чи пасовища на іншу (при цьому на старому місці залишаються "осілі" особини, а втрата поновлюється за рахунок перебігання інших полівок у протилежному напрямку); в) розліт пов'язаних із сушею птахів на сусідні острови архіпелагу в пошуках
73
належних умов існування (або поширення тлі по змішаному деревостану і різнотрав'ю в пошуках потрібної рослини).
М.Бігон, Дж.Харпер і К.Таунсенд звертають особливу увагу на проблему демографічного значення розселення, на фактори, які стримують ці процеси. Коротко зупинимося на них, особливо на окремих цікавих прикладах. Що таке розселення - втеча чи відкриття? Передусім розселення полягає в тому, то особини залишають безпосереднє оточення своїх родичів і сусідів, розріджуючи скупчення і вирішуючи тим самим проблему перенаселення. Особина немовби втікає у пошуках нового місця. Однак, як наголошують автори, це не просто "втеча", але одночасно і пошук, і, врешті-решт, відкриття. Розрізняють два типи такого "пошукового" розселення: 1) коли особини відвідують і "обстежують" інші ділянки, а потім повертаються в одну з них (зрозуміло, найпридатнішу) і там залишаються назавжди; 2) коли особини відвідують одне місце за іншим, а потім просто зупиняються, не "повертаючись" на старе. В межах останнього типу розселення розрізняють випадки, коли зупинки підвладні і не підвладні переселенню. Наприклад, поширення насіння рослин має "дослідницький" характер, хоча від самого насіння зовсім не залежить, потрапить воно чи не потрапить в сприятливі умови. Розселення ж тварин підпадає під будь-яку з наведених вище категорій, причому вони добре знають, на якому придатному варіанті їм зупинитися. Наприклад, тля (навіть крилата) не може протистояти панівному вітру, проте злетіти з того місця, де вона народилась, або ж випасти з повітряного потоку вона може. Тля перелітає на невеликі відстані, а отже, "відкриває" кращі місця, образно кажучи, зі своєї волі.
Пасивне розселення сушею і повітрям спостерігається у вигляді "насіннєвого дощу". Повітряний простір з його течіями є простором пасивного переміщення насіння багатьох видів рослин. Однак не лише крилатки клена підвладні повітряним потокам: восени перелітають на значні відстані павуки, тримаючись за своє павутиння, як за стропи парашута. Лісівники, знаючи напрям панівних вітрів, нарізають лісосіки з таким розрахунком, щоб вітер міг занести на зруби насіння сосни чи бука. Справді, на землю з повітря падає насіннєвий дощ, несучи нове життя в нові місцезростання.
В природі поширене розселення "силоміць" за допомогою активного носія. Насіння багатьох трав'яних рослин, які ростуть у нижніх ярусах лісу, мають колючки або шпичаки (гачки), що підвищує вірогідність пасивного поширення насіння ссавцями, які переносять його за допомогою шерсті. Відомими транспортувальниками насіння є птахи, які поїдають привабливі соковиті плоди, розсівають потім насіння разом зі своїм послідом.
74
Пасивними способами розселення користуються і окремі тварини. Наприклад, кліщі, як правило, сідають на молодого жука, який щойно вийшов з лялечки, і злазять з нього тоді, коли той перебрався на нову купу гною. Пасивне розселення течією води поширене в основному в комах, личинки яких переносяться товщею води. Таке розселення називають ще "дрейфом".
Демографічне значення розселення, як важливого демографічного процесу, заслуговує на підвищену увагу екологів. Наприклад, інтенсивне розселення колорадського жука в Європі в середині нашого століття завдало значної шкоди сільському господарству.
Спостерігається активне й пасивне занесення тварин людьми із найрізноманітніших кліматичних зон у міські острівні місцезростання (склади, теплиці, будинки, культури кормових рослин, озеленення).
На відміну від рослин, точний час завезення і походження тварин не завжди відомий. Давно, наприклад, оселився в Європі американський тарган Рeriplanetа атеrісапа. Лише в 1959 р. в Центральній Європі помітили завезеного з Японії жука-водолюба Сryptopleurum sultilae, який, напевно, зараз поширюється.
Одним із способів розселення є інвазія, або експансія виду, тобто
заселення особинами певного виду нових територій, які не були до цього зайняті його популяцією. Це розширення ареалу, зайнятого видом. Прикладом інвазії можуть бути спалахи хвороб, таких, як грип. У 1918р. ця хвороба, яка переносилась вірусом грипу, охопила всю Європу, обминувши лише Гренландію і Лабрадор, забравши понад 10 млн. життів, тобто більше, ніж п'ятирічна перша світова війна.
Виділяють два джерела екологічної інвазії: перше – континентальна ізоляція з часів крейдяного періоду, яка унеможливила оптимальне використання екологічних умов певного біологічного виду; друге – інтенсивний розвиток комунікацій на планеті.
2.5 Взаємодія організмів у середині популяції і за її межами
Динамізм популяції, який е запорукою життєдіяльності і життєздатності організмів, проявляється у взаємодії окремих особин як всередині власної популяції, так і з особинами інших популяцій. Розглянемо чотири процеси, які включають взаємодію між організмами, породжуючи природний відбір: конкуренцію, хижацтво, паразитизм і мутуалізм.
75
2.5.1 Внутрішньовидова конкуренція
Розрізняють чотири основні ознаки внутрішньовидової конкуренції. По-перше, при внутрішньовидовій конкуренції ресурс, за який особини борються, має бути обмеженим. У зв'язку з цим створюється ситуація для експлуатаційної й інтерфєрентної конкуренції, Перша полягає в тому, що всі особини одночасно експлуатують ресурси, але кожна з них використовує те, що залишилося від конкурента. У другому випадку одна особина не дає іншій зайняти існуюче місцезростання і використати її ресурси.
Першу форму конкуренції називають жорсткою, а другу – суперництвом. Приклад жорсткої конкуренції – це експеримент із зеленою падальною мухою, коли при досягненні певної граничної щільності особин культура повністю гинула. Суперництво можна проілюструвати на прикладі закладки гнізд: на наявні 100 дупел, які є в лісі, претендує 200 пар дуплогніздувальних птахів. Отже, сто пар птахів не зможуть влаштувати гнізд і не дадуть потомства. Другий можливий варіант – еміграція на іншу територію. Цю форму конкуренції називають ще ідеальною або безкомпромісною. По-друге, конкуруючі особини одного і того ж виду не є рівноцінними. Наприклад, сильний ранній росток може затіняти низькорослий екземпляр, який з'явився раніше.
По-третє, вплив конкуренції на будь-яку особину тим сильніший, чим тісніша взаємодія конкурентів. Отже, слід вважати, що наслідки внутрішньовидової конкуренції залежать від щільності.
По-четверте, кінцевим наслідком конкуренції є зменшення вкладу особини в наступне покоління, тобто порівняно з тим вкладом, який міг би бути зроблений за відсутності конкурентів. Виходячи з цього положення будують прогностичні моделі розвитку популяції.
Всі ці ознаки внутрішньовидової конкуренції, виявлені в процесі довготривалих спостережень в природі і лабораторіях, підтверджують думку Ч.Дарвіна про гостроту боротьби за існування, оскільки вона не тільки постійно загрожує життю багатьох особин, але й призводить до повної загибелі популяції через загальне пригнічення усіх особин.
Для того щоб зрозуміти складний механізм внутрішньовидової конкуренції, доводиться вивчати умови середовища і його ресурси, які є гарантом довготривалого і благополучного існування популяції, зростання її продуктивності та постійного відтворення. А тому йдеться про вивчення екологічної ніші популяції.
Термін екологічна ніша був уперше вжитий американцем Д.Гріннелом у 1917 р. при характеристиці місцезростання пташки пересмішника (Тохоstота redivirит) – мешканця заростей чагарника чаппаралі. Згодом у 1933 р. його співвітчизник Ч.Елтон дав нове визначення цього поняття:
екологічна ніша являє собою не лише певні умови середовища, але й спосіб
76
життя і спосіб добування їжі. За образним висловлюванням Ю.Одума (1975), місцезростання – "адреса" організму, а екологічна ніша – його "професія".
В 1952 р. англійський еколог Дж.Хатчинсон увів поняття багатовимірної ніші, під якою розумів систему з багатьма координатами екологічного простору в екосистемі, де проживає і відтворює себе особина (або популяція). Американський еколог Е.Піанка (1981) визначає
екологічну нішу як загальну суму адаптацій організму до певного середовища. Як і у випадку терміна "середовище", ми можемо говорити про нішу особини, популяції або виду. Різниця між оточуючим середовищем і нішою організму полягає в тому, що цей термін відбиває здатність організму засвоювати своє середовище і вказує шляхи його фактичного використання.
Тривале вивчення і аналіз екологічних ніш різних популяцій дали змогу виділити два типи – фундаментальний (передконкурентний або потенційний) і реалізований (постконкурентний або фактичний).
Розрахунок багатовимірної ніші вимагає великої кількості даних, які в природних умовах, враховуючи їх взаємозалежність, полікоординатність, нелегко одержати. Тому в екології часто використовують часткові ніші, в яких беруть до уваги лише окремі фактори середовища (наприклад, у відомій едафічній сітці П.С.Погребняка родючість ґрунту і його вологість). Розрізняють трофічні, едафічні, гідрологічні, математичні та інші види ніш. В одних організмів (стеноеків) вони вузькі (вільха чорна вимагає не лише більш родючих і зволожених ґрунтів, але й проточної води), в інших (евріеків) – широкі (сосна, яка трапляється як піонерний вид на бідних дюнних пісках і в багатих едафічних умовах судібров).
Поняття екологічної ніші стосується не лише популяції, але й конкретного організму. Це особливо видно на прикладі впливу щільності популяції на її динаміку, що часто спостерігають лісівники. Надмірна густота самосіву під пологом насадження чи сіянців у розсаднику призводить до того, що окремим особинам, особливо генетично слабшим або таким, що ростуть у дещо гірших умовах, не вистачає простору, поживних речовин чи світла.
Зазначимо, що кожна рослина для нормального забезпечення потреб розвитку вимагає певного ростового простору, величина якого залежить від виду, віку рослини і якості умов місцезростання. А.К.Каєндер ще у 20- х роках з'ясував, що сосна у фінських лісах у 50 років вимагає 2 м2, у 75 – 3,8 м2, у 124 – 11 м2, а у віці 150 років – 15 м2 простору росту. В умовах більш суворого клімату цей самий вид вимагає більшого простору, ніж у м'якшому. Надмірне загущення рослин зумовлює гостру внутрішньовидову конкуренцію, що відбивається в цілому на рості і розвитку дерев і призводить до можливого випадання з популяції особин, які виявляються нежиттєздатними.
77
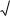
Внутрішньовидову конкуренцію особин можна розрахувати за допомогою рівняння Йоди, де середню масу і площу на одну особину
знаходимо таким чином: |
|
|
|
|||
W=Cd3/2=C/d3/2 d=1/a , |
(2.7) |
|||||
у =wd= |
C |
|
, |
(2.8) |
||
|
|
|
||||
d |
||||||
|
|
|
|
|
||
де а – середня величина, яка припадає на одну віддаль, d – щільність, у – середня маса на одиницю площі, w – середня маса рослини, С – константа.
Для видів, які належать до експлуатаційного типу конкуренції або конкуренції захоплення (scramble competition), наслідки зростання чисельності популяції можуть зумовлювати збільшення смертності через зупинення росту особин. Оскільки деякі тварини у дорослому віці мають фіксовані розміри тіла (туфелька), то для них ємність середовища може впливати головним чином на кількість особин, а не на їх розміри.
Розмір тварин, якщо вони мають невизначений ріст, який не зупиняється ще й у віці статевої зрілості (риби), визначається запасами корму. Якщо в перший ставок – говорить Р.Уіттекер, – випустити 100 мальків, а в другий 50, то розміри риби в першому можуть бути невеликими, тоді як у другому – у два рази більшими. Однак загальна маса риби може бути майже однаковою в обох ставках. Розміри рослин ще більше пристосовані до конкуренції за ресурси, які вони споживають. Із двох дерев одного виду і віку одне, яке росло весь час в затінку, у 100 років може бути завтовшки 5 см, а те, що виросло в умовах достатнього світла, – 50 см.
Здатність рослин змінювати розміри, – підкреслює Р.Уіттекер, - створює вірогідність парадоксальних наслідків конкуренції – зменшення щільності популяції зі збільшенням ресурсу середовища. Наприклад, у теплиці на двох ділянках однакової площі висіяно однакову кількість насіння. Перша ділянка отримує добрив у два рази більше, ніж друга. Рослини на краще удобреній ділянці ростуть швидше і мають більші розміри, а найрозвинутіші серед них особини виявляються сильними конкурентами (за світло, воду, поживні речовини) для тих, що ростуть повільно. Смертність при цій конкуренції захоплення висока, і до стадії зрілості доживає лише невелика кількість крупних рослин. Щільність і середня маса рослини пов'язані між собою експоненційною або логарифмічною, залежністю. Середня площа, яка припадає на одну рослину (і обернено пропорційна щільності), змінюється як квадрат лінійних розмірів.
Внутрівидова конкуренція має чимало негативних наслідків: вона не лише збіднює ресурси, що супроводжується затримкою в рості або
78
збільшенням смертності, але й призводить до токсикації (інтоксикації) навколишнього середовища, самоагресії або канібалізму, соціальної та репродуктивної нездатності. В цій ситуації популяція може збільшуватися доти, доки її чисельність не стає надмірною.
2.5.2 Міжвидова конкуренція
Еволюція кожної окремої популяції відбувалася у взаємодії з іншими популяціями, з якими вони утворювали певні угруповання. Одновидове угруповання може існувати лише в ідеальній ізоляції від зовнішнього світу і, мабуть, не довго. Життєвий потенціал виду, який дожив до наших днів, сформувався в процесі тривалої міжвидової боротьби за існування. Конкурентні стосунки є одним з найважливіших механізмів регулювання видового складу кожного угруповання, просторового розміщення видів та їх чисельності.
Американські вчені Е.Піанка (1981), А.Лотка (1922) і В.Вольтерра (1926, 1931) були першими, хто розробив порівняно міцну, хоч і дуже спрощену, теоретичну основу вивчення конкуренції. Їхнє рівняння конкуренції сильно вплинуло на розвиток сучасної екологічної теорії і є позитивним прикладом математичної моделі важливого екологічного явища. Крім того, це рівняння дало поштовх розробці декількох інших дуже корисних понять, які концептуально з ними не пов'язані (поняття коефіцієнтів конкуренції, матриць угруповань і дифузної конкуренції).
Розглянемо рівняння конкуренції Лотки – Вольтерра. Це, по суті, видозмінене логістичне рівняння Верхлюста – Пірла, причому обидва базуються на однакових припущеннях. Проаналізуємо це рівняння.
Конкурують два види, які характеризуються величинами щільності N1 N2 і мають в разі відсутності один одного граничну щільність насиченості К1 або К2. Логістичне рівняння для виразу змін щільності (N) у часі:
dN |
K N |
|
||
|
rN |
|
. |
(2.9) |
|
|
|||
dT |
|
K |
|
|
Нехай кожен з видів 1 і 2 мають свою швидкість росту (r1 граничну чисельність (k1 і k2), а коефіцієнти конкуренції (α1 характеризують вплив одного виду на інший, тоді:
dN |
|
|
r1N1 |
|
K |
N a N |
, |
||||||||
1 |
|
|
1 |
|
|
1 |
1 |
|
|||||||
dt |
|
|
|
|
|
|
|
K1 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|||||
dN |
|
|
r2 N |
2 |
K |
N |
|
a N |
|
|
|||||
|
2 |
|
|
2 |
2 |
|
2 |
|
2 |
. |
|||||
dt |
|
|
|
|
|
|
K2 |
|
|
|
|
||||
|
|
|
|
|
|
|
|
|
|
||||||
іr2) і
іα2)
(2.10)
(2.11)
79
Результати кожної реакції визначаються значенням коефіцієнта
конкуренції і граничної чисельності: |
|
||
αj <k j / k │ |
α│>k│/ kj |
(виживає тільки вид (2.10)); |
(2.12) |
αj >k j / k │ |
α│<k│/ kj |
(виживає тільки вид (2.11)); |
(2.13) |
αj >k j / k │ |
α│>k│/ kj (виживає або перший або другий вид); |
(2.14) |
|
αj <k j / k │ |
α│<k│/ k j |
(виживають обидва види) |
(2.15) |
Як бачимо, рівняння не приводять до єдиного висновку і свідчать, що різні наслідки конкуренції залежать від співвідношень α1,α2, k1, k2. Із чотирьох можливих варіантів три передбачають, що один із видів буде згасати. Останній випадок (2.15) цікавіший: два види можуть вижити при сумісному урівноваженому існуванні. Це трапляється у випадку роздільного вирощування видів у культурі, де один вид лімітується лише своєю власною внутрішньовидовою конкуренцією в одній частині культури, а інший, поряд, своєю (наприклад, шкілка дерев чи чагарників). У цьому випадку можлива стійка конкуренція, оскільки види поділяють простір культури між собою таким чином, що кожен з них вдало конкурує у своїй власній частині культури.
Між рослинами і тваринами існують дві відміни, які мають велике значення для вивчення конкуренції. По-перше, лише в незначної кількості рослин період генерації триває менше року, тому у багатьох випадках екологи рослин не мають можливості здійснювати тривалі експерименти, які б дали змогу продемонструвати конкурентне виключення. По-друге, на ріст і виживання рослин великий вплив мають різноманітні умови, в яких вони живуть. Наприклад, в умовах великої скупченості ріст рослин сповільнюється, і вони не досягають повного розвитку, хоч і можуть продукувати насіння. На відміну від цього популяції тварин звичайно відповідають на перенаселеність підвищенням смертності і затримкою росту.
Основою для спостережень конкурентних стосунків можуть служити три пробні моделі, які описують: 1) недосконалу конкуренцію, коли міжвидова конкуренція є лімітуючим фактором, але не веде до повної елімінації (усунення) одного з конкурентів з арени взаємодії; 2) досконалу конкуренцію, описану моделями Гаузе і Лотки – Вольтерра, коли один вид поступово елімінується в процесі конкуренції за загальний ресурс; 3) наддосконалу конкуренцію, коли ефект подавлення дуже сильний і проявляється негайно, наприклад, під час виділення антибіотиків (алелопатія). Наочним прикладом такої "надсильної" конкуренції може бути також хижацтво.
Для кращого розуміння міжвидової конкуренції варто зупинитися на таких поняттях, як співіснування і конкурентне виключення, екологічне заміщення видів, екологічна компресія і вивільнення, співіснування і розподіл ресурсів, а також еволюційна дивергенція.
80
Співіснування і конкурентне виключення – одне з надзвичайно цікавих і слабо вивчених екологічних явищ. Дослідження їх в польових умовах і в лабораторії дають протилежні дані про природу. Спостерігаючи за життям рослинного і тваринного світу, ми частіше стаємо свідками того, як співіснують види, а не як вони борються за існування. На Шацьких озерах, що на Волині, поряд плавають зі своїми виводками кілька видів крижнів, дикі гуси, лебеді, які живляться рибою. В свіжій грабовій бучині на Розточчі поблизу Львова живуть поряд, співіснуючи, 19 видів дерев, 24 чагарників і чагарничків, 72 трав'яних рослин. Насправді це далеко не так: конкурентна боротьба за використання ресурсів, а отже, за існування точиться безперервно, але в природі вона не так помітна, як в умовах лабораторії.
Г.Ф.Гаузе першому вдалося в лабораторії створити умови для співіснування двох схожих видів, правда, використовуючи одне і те ж поживне середовище. Культуру двох видів туфельок вирощували в двох варіантах: види жили порізно і сумісно. В першому випадку перша і друга туфельки (Раrатесіит аurelіа, Р.саиdаtит, P.bursaria)
розмножувалися і підтримували постійну чисельність популяції.
Згодом аналогічні дослідження Г.Ф.Гаузе здійснив і з борошняним хрущиком (Tribolium). Ці дрібні жуки весь свій життєвий цикл можуть завершити у банці з борошном, яка служить для них одночасно і місцем зростання, і кормом для личинок та імаго. Коли в це однорідне середовище помістили два різних види хрущика, то виявилося, що один з них переміг і успішно розвивається, витіснивши іншого.
Результати лабораторних експериментів з конкуренції привели до формулювання принципу конкурентного виключення, який ще називають законом Гаузе: два види не можуть співіснувати, якщо вони залежать від одного і того ж лімітуючого середовища. Підкреслюємо – лімітуючого середовища, оскільки лише ті ресурси, які лімітують ріст популяції, можуть створювати основу конкуренції.
Екологічне заміщення видів. Приклад з мучним хрущиком дає можливість простежити ще одне цікаве явище – екологічне заміщення видів. Коли два види цього жука (T. costaneum і T.coninsum) вирощувати разом у пшеничному борошні, то при невисокій температурі і низькій вологості T. costaneum витісняє T.coninsum, а в теплому і вологому середовищі T. costaneum сам виявляється витісненим. Отже, вид може проявити конкурентну здатність в субоптимальних умовах; наслідок конкуренції не залежить від того, як функціонує даний вид в умовах відсутності конкуренції, а визначається відносною продуктивністю видів при сумісному вирощуванні. На користь такого висновку свідчить той факт, що вид, який перемагав у конкуренції при будь-яких поєднаннях температури і вологості, не завжди виявлявся найпродуктивнішим, коли його вирощувати в ізоляції при тих же умовах.
81
Конкуренція пов'язана зі специфічною взаємодією між видами, яка рідко проявляється при окремому спостереженні за кожним з них. Прикладом цього явища може служити сумісне і роздільне зростання двох видів дуба – звичайного (Quercus robur) та скельного (Q. petraea). В свіжих типах можна побачити ці два види поряд, в сухих типах, особливо з кам'янистою підстилаючою породою, дуб звичайний заміщається дубом скельним.
Екологічне вивільнення й екологічна компресія – явища, протилежні за своїм змістом. Екологічне вивільнення полягає в усуненні конкурента і одержанні таким чином додаткових ресурсів. Чимало прикладів екологічного вивільнення одержано лісівниками, які вивчали вплив рубок догляду на формування якісної деревини. Вилучаючи особини, які відстали в рості, а також "небажані" види, створюються сприятливі умови (освітлення, вологості, мінеральної поживи) для "бажаних" видів.
Екологічна компресія зумовлена введенням конкурента. Явища екологічної компресії часто спостерігаються на віддалених від материка островах з їх обмеженим видовим складом як рослин, так і тварин. Коли сюди потрапляють види, які були витіснені з материка, вони швидко адаптуються до нових умов місцезростання з незначною різноманітністю конкурентів і швидко поширюються (кролі і кактуси в Австралії).
Співіснування і розподіл ресурсів. У попередніх варіантах конкуренція розглядалася як виключення і успіх, елімінація і виживання, переваги і подавлення. Цими термінами описуються процеси, які спостерігались в угрупованнях у минулому, тоді як співіснування – стан, який маємо сьогодні. Упродовж десятиріч екологи вивчають умови, необхідні для співіснування видів. Математичний аналіз міжвидової конкуренції свідчить, що якщо даний вид обмежує своєю чисельністю популяції іншого виду, і навпаки, то між такими двома видами можливе співіснування. Ці умови виконуються у тому випадку, якщо кожний вид використовує дещо інший ресурс, ніж інший. Відомо, що види уникають екологічного перекриття, розподіляючи між собою наявні ресурси відповідно до їх розмірів і форми, хімічного складу, місця, де вони трапляються, а також їх сезонності.
Як бачимо, наслідок конкуренції значною мірою залежить від того, як конкуруючі види використовують (вдало чи невдало) надзвичайно неоднорідне середовище, яке переважно складається з окремих ділянок ("плям") зі сприятливими і несприятливими умовами. Стійкість до несприятливих умов дає можливість окремим видам знаходити поживу в той час, коли інші гинуть. Конкуруючі види, як правило, не зустрічаються в одному місцезростанні і поділяють між собою не лише кормові ресурси, але й простір. Наприклад, кожен із п'яти видів американських славок (Dепdrоіса), які гніздяться в американському штаті Мен, годується на
82