

Ekologiya
.pdfДобова динаміка залежить від освітлення, температури, складу повітря, його руху, зволоження і т.п. Коливання добового режиму особливо помітні в угрупованнях рослин з явними ознаками фототаксису
– руху організмів під впливом однобічного освітлення: позитивний – до джерела освітлення, негативний – від джерела освітлення. Наприклад, це повороти суцвіть рослин, які спостерігаються на плантаціях соняшнику. (Наслідком фототаксису є веретеноподібна форма гілок і стовбура дуба звичайного).
Сезонні або фенологічн, зміни зумовлені зміною пори року. В хвойних ценозах вони ледь помітні, тоді як у листяних лісових фітоценозах яскраво виражені (наприклад, сезонна зміна в березняку). В дібровах виділяють близько 10 фенофаз.
Різнолітня динаміка залежить головним чином від метеорологічних умов року (наприклад, посуха, дощове літо і т.п.).
Стадійна динаміка. В природі часто спостерігаються і незворотні зміни. Вони зумовлюють зміну або сукцесію одних угруповань іншими на одному і тому ж місці. Ці зміни, або сукцесії, можуть бути природними або ж антропогенними. В першому й другому випадках лісівники та паркобудівники повинні добре уявляти розвиток фітоценозу як у часі, так і в просторі.
3.6.3 Систематика та класифікація фітоценозів
Природа багата на фітоценози. З давніх часів людина навчилась розрізняти луки, ліси, степи, болота. Ліси підрозділяють на листяні (дубові, букові), хвойні (ялинові, ялицеві, соснові) та мішані. Завдання еколога полягає у виявленні та класифікації всіх фітоценозів за допомогою угруповань.
Кожна наука під час систематизації об'єктів, що вивчаються, застосовує певні одиниці. Наприклад, при систематиці рослин, як і тварин, використовують такі систематичні одиниці або таксони: вид, рід, родина і т.п. У фітоценології фітоценоз вважають аналогом поняття "рослина", тобто фітоценоз має загальне значення як закономірне поєднання взаємозв'язаних рослин і, відповідно, не має таксономічного рангу. Ділянки рослинності об'єднуються в нижчу одиницю систематики фітоценозів – асоціацію. Більшість українських, російських, скандинавських і американських вчених-фітоценологів визначають асоціацію за домінуючими видами.
Асоціація – це сукупність ділянок рослинності з однорідною фізіономічністю та спільними домінантами у всіх ярусах. Таке визначення відбиває суттєві риси асоціації: найважливіші види, їх покриття, ярусність, зовнішність, зв'язок із зовнішніми умовами (частково).
123
Тривалі пошуки підходів до систематики як природних, так і культурних або штучних насаджень дали змогу виділити три основні напрями в класифікації насаджень: 1) типологічний – в основу покладені типи насаджень; 2) антропогенний – за рівнем окультуреності рослинності; 3) антропогенно-фітоценотичний – за рівнем окультуреності фітоценозу.
Типологічний напрям зосередив у собі декілька класифікаційних систем, зокрема:
екологічну (Морозов та українська школа лісівників Олексіїва-По- гребняка);
фітоценологічну (Сукачов та московська-ленінградська школа лісівників);
генетичну (Колесников та сибірська школа лісівників);
еколого-фітоценотичну (Голубець, Малиновський та карпатська школа лісівників);
багатофакторну (Бєлов, Рисін).
На практиці, особливо в умовах сильного антропогенного впливу на зелені насадження, потрібна така класифікація, яка б сприяла раціональнішому веденню господарства, зокрема лісопаркового. Згідно з В.М.Сукачовим, тип лісу слід розглядати не як господарську, а як природно-історичну одиницю. Планувати господарство необхідно за допомогою типу лісу (Морозов).
Якщо господарська одиниця лісів третьої групи (експлуатаційних) може включати декілька типів лісу, то в насадженнях першої групи, наприклад у лісопарковій госпчастині, господарські заходи слід планувати на парцелярному рівні: група сосен чи барвінкова галявина вимагають індивідуального підходу.
На кафедрі екології та ландшафтної архітектури Українського державного лісотехнічного університету в 70-х роках (тоді кафедра лісівництва) розроблена класифікація насаджень зелених зон міста (Кучерявий, 1973). В її основу були покладені елементи екологофітоценотичної (Голубець, Малиновський), антропогенної (Хохлов, Фурсаєв) та антропогенно-фітоценотичної (Ніценко) класифікацій.
Елементарною одиницею (нижчим таксоном) цієї антропо-еколого-фі- тоценотичної класифікації є асоціація. Асоціація (від латин, associo – з'єдную, поєдную) – рослинне угруповання, яке характеризується однорідним флористичним складом, наявністю провідних (домінантних) видів, певним кількісним співвідношенням між окремими видами, а також ярусністю, послідовністю фенологічного розвитку рослин протягом вегетаційного періоду та продуктивністю рослинної маси. Найвищим таксоном є тип рослинності (наприклад, деревна чи лучна рослинність), далі формація (букові ліси), субформація (грабово-букові ліси), екологічна група асоціацій або тип лісу (свіжа грабова бучина), асоціація (природна чи похідна). Похідні асоціації, в свою чергу, діляться на природні
124
(саморегульовані) і штучні (регульовані), а останні – на утворені в слабозмінених, середньозмінених і сильнозмінених умовах місцезростання.
Штучно створені фітоценози називають культурфітоценозами. Найбільше таких фітоценозів створено на урбанізованих територіях: сильваценози (лісові), фрутоценози (чагарникові), помологоценози (садові), вітоценози (виноградники), флороценози (квітники), пратоценози (газони, луки), агроценози (поля із зерновими та просапними культурами), стрипоценози (захисні смуги), акваценози (декоративні ставки та басейни із водяною рослинністю).
Фітоценологи в останні роки приділяють значну увагу рудероценозам
– бур'яновим рослинним угрупованням, які займають значні урбанізовані території міст і селищ. Часто ці рудеральні фітоценози відіграють позитивну роль і вирізняються високою стійкістю до антропогенних навантажень. Наприклад, споришево-подорожникові асоціації в багатьох житлових мікрорайонах великих міст не лише відіграють значну естетичну роль, але й є стійкими до витоптування. Крім того, рудеральні фітоценози в урбогенних і техногенних умовах є екологічними нішами для багатьох видів фауни.
3.6.4Типи асоціацій
Уфітоценології та геоботаніці використовують два способи класифікацій асоціацій. Перший полягає у застосуванні подвійних латинських назв, наприклад, Ріпеtит hylocomiosum (бір зеленомошник), Fagetum asperulosum (бучина маренкова) тощо. Для правильної побудови подвійної латинської назви потрібно додати закінчення "etum" і "osum" до коренів слів, які визначають за їх родовим відмінком, відкидаючи закінчення, утворене цим відмінком. При необхідності точного уявлення про рослинне угруповання можна розширити найменування асоціації до трьох чи навіть чотирьох назв, наприклад: Betuletum mixto herbosum (березняк із мішаним трав'яним покривом) або Picetum sphagno-herbosum (смеречник сфагново-трав'яний).
Інший спосіб найменувань асоціацій полягає у переліченні найголовніших домінуючих видів, найчастіше за ярусами, наприклад, Аs.
Ріпиs silvestris – Vaccinium myrtillus + Нуlосотіит splendens (сосновий бір+чорничник із моховим покривом). Якщо ж в угрупованні яруси виражені чітко, на перше місце ставлять домінуючий вид, а на друге – субдомінуючий: Аs. НеІісhrysum аrепаrіит + Dianthus аrепаrius (цмин пісковий + гвоздика піскова). Однак на цьому піщаному березі, де ростуть зазначені оліготрофи-ксерофіти, зустрічаються асоціації з більшою кількістю більш-менш однаково представлених видів. Тоді їх назва буде включати необхідну для повного уявлення кількість видів: As. Коеlеrіа
125
glauca + Stipa joannis + НеІісhrysum аrепаrіит + Sedum acre + Косhia laniflora. Назву цієї асоціації можна записати і українською мовою: Ас. келерія сиза + ковила Іоанна + цмин пісковий + очиток їдкий + віниччя шерстистоквіткове.
Якщо перед назвою рослини стоїть знак "плюс", як це бачимо в попередніх назвах асоціацій, то це означає, що участь даного виду у складі угруповання є менша, ніж виду, який стоїть попереду. Якщо ж назви двох чи більше видів з'єднані знаком "тире", то це означає, що участь цих видів у складі угруповання приблизно однакова. Наприклад, у парку маємо угруповання із рівним складом дерев: Аs. Асеr рlatanoides – Асеr pseudoplatanus – Тilia соrdata – Веtula pubescens (Ас. клен гостролистий – клен-явір – липа дрібнолиста – береза пухнаста).
Для означення асоціацій використовують одночасно й обидва знаки ("–“ і "+"), зокрема, перший для опису домінантного, а другий – субдомінантного ярусів: Аз. Ріпиs silvestrisїгз – Quercus robur – Веtula verrucosa + Рteridium aquilinum + Риlтопаrіа апgиstifolia + Сrista саstrensis
(Ас. Сосна звичайна – дуб звичайний – береза бородавчаста + орляк звичайний + медунка вузьколиста + пір'ястий мох). Таке написання характерне для російсько-шведської геоботанічної школи. Американоанглійська школа користується в основному внесенням в назву асоціації лише родових назв: Ріпиs — Juniperus аss (сосново-ялицева ас.).
Найбільшого поширення набула класифікація, яку використовує швейцарська школа Браун-Бланке і яка полягає у переліченні переважаючих рослин: Рlапtаgini – Роlуgопаtит аvicularis аss.
(подорожниково-споришева ас.) або ж назві асоціації за переважаючим видом – Роlуgопит (подорожникова), Таrахасит (кульбабова) тощо.
Школа Браун-Бланке використовує систему найменувань як для асоціацій, так і для більш крупних таксономічних одиниць. Після найменування асоціацій чи іншої геоботанічної одиниці свого часу БраунБланке запропонував ставити прізвище дослідника, а деколи і рік, в якому ця одиниця була вперше описана. Наприклад, для газонів, засмічених бур'янистою рослинністю, характерна асоціація Lolio — Рlапtаgіпеtит (пажитнице-подорожникова). У повному таксономічному виразі її записують таким чином:
Клас |
Plantageneta majoris R.Tx.et Prsq. 1950 р. |
Рід |
Plantagenetalia R, Tх. 1947 р. |
Союз |
Роlудопіоп аvicularis Вr.Вl. 1931р. |
Асоціація |
Lolio – Рlапtаgіпеtит (Link, 1921) Воgег. 1930р. |
Якщо назва асоціацій закінчується на "еtит", то назва субасоціацій закінчується на "еtоsит", наприклад, асоціація (Querceto — Сарrinetum Тх.
1930, субасоціація (Querceto – Сарrinetum саrісоsит (ріlоsae) (дубово-
грабово-волосиста).
126
3.7 Біоценотична структура угруповань
3.7.1 Принципи функціонування біоценозу
Різноманітні залежності і зв'язки реалізуються в біоценозі окремими компонентами. Однак не всі типи структур мають істотне екологічне значення. Важливим є те, яку функціональну роль відіграє той чи інший компонент структури в основних біоценотичних процесах. Головним чином це стосується потоку речовини й енергії, тобто енергетичного господарства біоценозу.
Елементом функціональної структури біоценозу може бути як популяція виду, так і група популяцій, включених в обіг матерії й енергії даного біоценозу. Структура біоценозу адаптована до функціонування в кожних конкретних умовах середовища і забезпечує нормальне функціонування системи, виходячи з трьох принципів:
1)оптимізації біологічної продукції екосистеми;
2)забезпечення обігу матерії й енергії;
3)стабілізації процесів.
Перший принцип визнає основи біологічної продуктивності екосистеми і, зокрема, утворення первинної продукції. Сонячна енергія, акумульована автотрофними організмами у вигляді високоенергетичних органічних зв'язків, є основою процесів перетворення матерії усіх біологічних компонентів системи. Структура біоценозу на рівні як автотрофів, так і гетеротрофів повинна забезпечити в разі змін середовища оптимальну, тобто близьку до максимальної, первинну продукцію екосистеми .
Другий принцип стверджує, що кругообіг матерії й енергії є основною властивістю екосистеми. Матеріали, які кружляють у межах систем, передаються між біологічними компонентами та середовищем. Гальмування обігу матерії хоча б в одному пункті екосистеми може призвести до порушення системи й її дезінтеграції.
Третій принцип стосується процесів, що відбуваються між біотичними й абіотичними компонентами. Реалізація цих процесів є спонтанною. Ступінь регуляції екологічних процесів відбиває рівень еволюційного розвитку біоценозу, їх адаптації до умов середовища, в яких формується даний біоценоз.
Контроль продукції, а також кругообіг матерії і його стабілізація є основною функцією, яку виконує біоценотична структура екосистеми. Це завдання реалізується через структури різного рівня, які спираються на відмінні типи коакцій між біологічними компонентами.
Виділяють три типи структур, які накладаються одна на одну в межах біоценозу: 1) трофічна; 2) конкурентна; 3) паратрофічна.
127
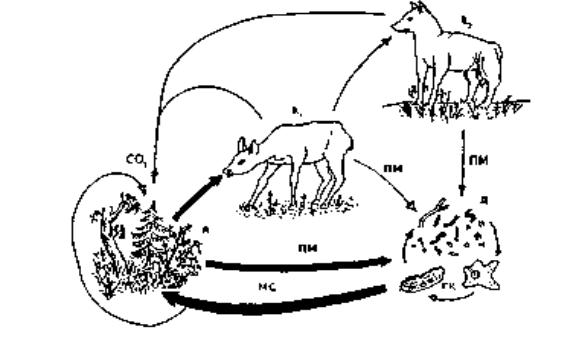
3.7.2 Трофічна структура біоценозів
Будь-яке угруповання можна представити у вигляді кормової мережі, яка являє собою схему всіх трофічних зв'язків між видами, що входять до складу цього угруповання. Кормова мережа звичайно складається із декількох ланцюгів живлення, кожний з яких є окремим її каналом
(рис3.1).
Рис 3.1 – Взаємозвязок продуцентів (п) з консументами 1-го порядку (К1), консументами 2-го порядку (К2) та деструкторами (д) і ґрунтовими консументами (ГК); ПМ – продукти матаболізму, мертвий випад; МС – мінеральні солі.
Завдяки кормовим взаємостосункам у біоценозах здійснюється трансформація біогенних речовин, акумуляція енергії і розподіл її між видами (популяціями). Чим багатший видовий склад біоценозу, тим різноманітніші напрями і швидкість потоку речовин і енергії. Ланцюги живлення або канали живлення, якими постійно перебігає енергія, прямо чи опосередковано об'єднують всі організми в єдиний комплекс.
Перший трофічний рівень представлений первинними продуцентами
або автотрофами. До них належать зелені рослини, які здатні використовувати сонячне проміння для утворення хімічних сполук, багатих на енергію. Первинні продуценти – це найважливіша частина біоценозу, тому що практично решта організмів, що входять до його складу, прямо чи опосередковано залежать від постачання енергією, якою запаслися рослини.
128
Крім первинних продуцентів до складу біоценозу входять гетеротрофи (від грецьк. гетеро – інший, трофе – корм) – організми, які використовують для споживання (корму) готові органічні речовини, представлені консументами і деструкторами. Перша група утворює
ланцюги поїдання, а друга – ланцюги розкладу.
Другий трофічний рівень утворюють травоїдні тварини, яких називають первинними консументами. М'ясоїдних, які живляться травоїдними, називають вторинними консументами або первинними хижаками; вони перебувають на третьому трофічному рівні.
Хижаки, які живляться первинними хижаками, в свою чергу, становлять четвертий трофічний рівень і називаються третинними консументами. Тварин, що споживають вторинних хижаків, називають третинними хижаками, або ж четвертинними консументами, і т.д.
Оскільки чимало тварин всеїдні (живляться як рослинами, так і тваринами), тобто одночасно одержують енергію з декількох різних трофічних рівнів, їх неможливо віднести до відповідного рівня. Звичайно вважають, що такі організми входять відразу до декількох трофічних рівнів, а їхня участь в кожному рівні пропорційна складу вжитої ними їжі.
Кінцеву ланку кормового ланцюга утворюють так звані деструктори або біоредуктори – організми, які розкладають органічні речовини. Це переважно мікроорганізми (бактерії, дріжджі, гриби-сапрофіти), які селяться на трупах і екскрементах і поступово їх руйнують. Завдяки їхній діяльності відбувається повернення в мінеральне царство елементів, які містяться в органічних речовинах.
Гриби, наприклад, в основному беруть участь у розкладі клітковини рослин, а бактерії розкладають трупи тварин. Мікроорганізмидеструктори виконують і інші функції: вони продукують інгібітори (наприклад, антибіотики) або, навпаки, речовини-стимулятори (наприклад, деякі вітаміни), екологічне значення яких дуже важливе, однак ще мало вивчене.
Серед кормових ланцюгів, які починаються з автотрофних рослин, можна передусім виділити ланцюги хижаків і ланцюги паразитів.
Кормові ланцюги хижаків ідуть від продуцентів до травоїдних, які поїдаються дрібними м'ясоїдними, останні, в свою чергу, поїдаються крупними хижаками і т.д. В міру пересування цим ланцюгом хижаків тварини в більшості випадків збільшуються розмірами і зменшуються чисельно.
Наприклад, відносно простий і короткий ланцюг має такий вигляд
(рис.3.2):
Трава – – > коник – – > просянка – – > вуж – – – > сова,
129
продуцент І консумент II консумент III консумент IV консумент; а більш довгий і складний ланцюг, який включає консументи п'ятого порядку, виглядає таким чином:
сосна звичайна — > попелиця — > сонечко — > павуки — > комахоїдні птахи — > хижі птахи.
Кормові ланцюги паразитів на відміну від попередніх ведуть до організмів, які зменшуються в розмірах і збільшуються чисельно:
|
Сонце |
|
Зелені рослини |
|
Первинні |
|
|
продуценти |
Рослиноїдні |
Комахи, |
Споживачі |
тварини |
травоїдні ссавці |
першого порядку |
|
тощо |
|
Хижаки { |
Комахоїдні птахи |
Споживачі |
|
другого порядку |
|
Плазуни |
Споживачі |
|
|
|
третього порядку |
|
Хижі птахи |
Споживачі |
|
|
четвертого порядку |
Рис. 3.2. – Спрощений наземний кормовий ланцюг, який показує послідовність трофічних рівнів
Кормові ланцюги можуть починатися не лише з автотрофних організмів (автотрофний тип), але і з неживих органічних речовин (детритний тип). Це спостерігається тоді, коли в ролі консументів виступають детритоїдні організми, які належать до різних системних груп (безхребетні, які живуть в ґрунті, харчуючись опалим листям або ж бактерії чи гриби, які розкладають органічні речовини). Тут спостерігається чітка узгодженість: тварини створюють умови мікроорганізмам, розкладаючи трупи загиблих тварин і рослин.
Кормові ланцюги, які починаються з рослиноїдних s детритоїдних організмів, найчастіше представлені в екосистемах одночасно, але майже завжди один з них домінує. Однак у підземній сфері, де існування хлорофільних організмів неможливе, оскільки туди не доходить сонячне світло, існують лише ланцюги, які починаються з детритоїдних форм.
Вивчення трофічних зв'язків свідчить, що в природі жодний живий організм не може існувати ізольовано. Більше того, живі організми в межах біогеоценозів або ж біогеоценотичних комплексів створюють спеціалізовані об'єднання – консорції. Характерною особливістю цих об'єднань є те, що вони звичайно формуються на базі популяцій
130
автотрофних рослин (сосна, смерека, береза, пшеничне чи картопляне поле тощо). Такі популяції називають детермінантами, а види, які об'єднуються довкола них, – консортами. Звичайно консорцію зображають у вигляді концентричної фігури з рослиною-ядром (детермінантом) та оточуючими його консортами, які, в свою чергу, утворюють консорти першого, другого, третього порядку і т.ін.
Слід звернути увагу на трофічні рівні кормових раціонів. Організми вважають такими, що належать до одного трофічного ріння в тому випадку, коли в кормовому ланцюгу вони віддалені від зелених продуцентів однаковим числом трофічних рівнів. Трофічний рівень і кормовий ланцюг не завжди мають струнку послідовну схему. Всеїдні тварини годуються одночасно рослинним і тваринним кормом, а деякі хижаки мають широкий набір жертв. Оскільки дана рослина чи тварина може служити кормом для різних як живоїдних, так і травоїдних, кормові ланцюги часто перетинаються один з одним, складаючи кормову мережу, або сітку. Наприклад, трава може бути спочатку спожита не кроликами, а іншими травоїдними; кролика замість лисиці може з'їсти орел і т.д.
Щоби встановити існування кормових мереж і трофічних рівнів, слід знати харчовий раціон різних видів. Для цього існують різні способи.
По-перше – це безпосереднє спостереження з використанням астрономічних телескопів, прихованих відеокамер тощо.
По-друге, це аналіз вмісту шлунків. Особливо цей метод використовують щодо птахів. Французький еколог Р.Дажо наводить приклад з вивченням вмісту шлунка сороки. В ньому, зокрема, виявилося 92 представники різних видів комах. У цілому в шлунку було 494 комахи, в тому числі жуків – 302, метеликів – 39, прямокрилих – 16, двокрилих – 14, коників – 10. Крім того, виявлено 112 павукоподібних, 35 ракоподібних, 4 молюски, 3 хребетні. Слід відзначити, що понад 80% – це жертви довжиною 0,6-1,5 см. Вибір проявляється також щодо біотопу: сорока в основному живилася на лужках.
А ось так виглядає раціон однієї пари яструба, кормове угіддя якої влітку досягає 3700 га, а взимку – 5000 га (поблизу Гамбурга, Німеччина): хижаки (домашні коти, горностаї, ласки, дрібні хижі птахи) – 1,02; воронові (ворона, сорока, сойка, галка) – 7,04; дрібна дичина (заєць, кролик, куріпка, фазан, голуб) – 43,04; водоплавні птахи (мандаринки, качки, лисухи) – 1,01; домашні птахи (кури, голуби) – 20,25; різні птахи – 27,20; дрібні ссавці – 0,24. Як бачимо, склад жертв яструба різноманітний: 65 видів ссавців і птахів, причому переважають голуби, куріпки, граки, воронові і кролики. Метод аналізу вмісту шлунка має свої недоліки, оскільки у шлунках трапляються окремі м'які частинки тіла, походження яких важко встановити.
У вивченні кормових раціонів широко використовують радіоізотопний метод. Рослини, які поїдаються певного групою тварин,
131
поливають розчином ізотопного фосфору (Р32). Розчин швидко поглинається тваринами. Через рівні відрізки часу відловлюють на досліджуваній ділянці тварин і вимірюють їх радіоактивність. Виявляється, що тварини, які активно живляться цими рослинами, тобто своїм основним раціоном, швидше набувають радіоактивності. Тварини, які живляться в основному іншими рослинами, стають радіоактивними значно пізніше – через 2-5 тижнів. У павуків вона з'являється через п'ять тижнів, а у равликів і жужелиць майже не спостерігається, тобто вони майже не вживають досліджуваних рослин.
Трофічну структуру можна зобразити графічно у вигляді екологічних пірамід, основою яких служить перший рівень продуцентів, а наступні рівні утворюють сходинки і вершину піраміди. Екологічні піраміди відносять до трьох типів: 1) піраміди чисельності; 2) піраміди біомаси; 3) піраміди енергії.
Піраміда чисельності. Якщо представити кормовий ланцюг хижаків у вигляді прямокутників однакової висоти, розташованих один на одному, довжина яких пропорційна кількості особин у кожному трофічному рівні (особин на 1м2 в рік), то отримаємо фігуру, яку називають пірамідою чисельності. Вона тим вища, чим більша кількість трофічних рівнів включає даний ланцюг. Оскільки кількість особин від першого трофічного рівня до останнього звичайно зменшується, піраміда має форму трикутника, оберненого вершиною догори.
У кожній екосистемі дрібні тварини кількісно перевищують і розмножуються швидше. Наприклад, ґрунтова фауна за розмірами тіла розміщується таким чином: 0-1 мм - 4500 екз., 1-2 - 2000, 2-3 - 600, 3-4 - 200, 4-5 мм - 300 екз.
Для кожної хижої тварини існує нижня і верхня межі розмірів їхніх жертв. Верхня межа визначається тим, що хижак не в стані подолати тварину, яка перевищує розміри його власного тіла. Нижня межа визначається тим, що надто малий розмір здобичі робить полювання на неї нераціональним. Дрібну здобич хижаку довелося б ловити в таких кількостях, що це виявилося б неможливим або внаслідок її нестачі або ж через обмеженість часу. Тому для хижаків кожного виду, як правило, існує оптимальний розмір жертви. Елтон наводить такий приклад: для підтримки життя одного лева необхідно 50 зебр на рік; кулик-сорока, що селиться на Фолклендських островах (поблизу Аргентини), живиться молюсками, розмір яких перевищує 45 мм, тоді як дрібніші особини від нього вислизають; муха цеце (озеро Вікторія) нападає на ссавців і птахів, еритроцити яких мають діаметр 7-8 мк, але не чіпає рибу протоптерус, оскільки її еритроцити діаметром 41 мк надто великі, щоб пройти через канал хоботка мухи.
В кормовому ланцюзі спостерігається збільшення розмірів і зменшення чисельності особин при переході від трофічного рівня п до
132