

Ekologiya
.pdfРис. 2.4 - Деякі особливості форми кривих росту популяції при
зображенні в лінійному масштабі: А – J-подібна крива (експоненціальний ріст); Б
– S-подібна крива (сигмоїдальний ріст) і деякі їх варіанти.
63
Вона відбиває існування максимальної щільності популяції і, враховуючи можливе віддзеркалення сигмоподібної кривої (такий же плавний спад), має вигляд гіперболи (тому цей тип кривої часто називають ще гіперболічним) (рис.2.3,а,б).
В 1925 р. ця крива вдруге була відкрита Пьорлем, який використовував її для пояснення росту будь-якої популяції тварин, що має обмежені, але все ж поповнювані запаси їжі. Рівняння логістичної кривої (швидкості росту) має такий вигляд:
dN |
K N |
|
|
||
|
rN |
|
|
, |
(2.4) |
|
|
||||
dt |
|
K |
|
|
|
де N – чисельність (щільність) популяції; К – максимальна кількість особин, здатна жити у даному середовищі, тобто відтворити асимптоту кривої; dN/dt – коефіцієнт росту. Наприклад, у випадку популяції інфузорії з вихідною чисельністю 100 особин і з 200 особинами через 1 год коефіцієнт росту дорівнює 100/1=100/год, а коефіцієнт росту на особину dN/Ndt = 100/100-1=1 на особину за 1 год. Аналогічно визначають коефіцієнти народжуваності (b=dN/dt ) і смертності (d=dN/dt). Різниця r=b-d являє собою коефіцієнт приросту (стосується лише ізольованої популяції, де немає ні еміграції, ні імміграції).
Крива у верхній частині (рис.2.4) має S-подібну (сигмоподібну) форму. В популяції сигмовидного росту розрізняють три фази (навіть якщо вони пов'язані переходом одна з одною). У ранній фазі популяція майже не залежить від корму та простору і росте зі швидкістю, близькою до значення rN, що показано чіткіше через логарифмічну шкалу. У середній фазі проявляється лімітуючий вплив оточуючого середовища, відбувається сповільнення росту внаслідок збільшення смертності (або зменшення народжуваност, або з обох цих причин одночасно). В останній фазі популяція досягає своєї асимптоти і стабілізується близько своєї допустимої чисельності К.
Експоненціальна крива виражає "біотичний потенціал" - здатність до розмноження. Вченими встановлено, що через ослаблення біотичного потенціалу на нашій планеті вимерло 500 млн. біологічних видів (тепер у біосфері налічується 1,8 млн. видів). Перша спроба розкрити поняття біотичного потенціалу належить англійському вченому Чепману, який на початку 30-х років визначив його як природну здатність видів збільшувати чисельність. Записується він як показникова функція виду:
Nt = rT , |
(2.5) |
64
де Nt – кількість потомства через Т поколінь; r – кількість потомства від однієї особини впродовж покоління.
Циклічний тип – характеризує динаміку чисельності популяцій, яка регулярно повторюється під впливом циклічних змін у середовищі його існування (коливання температури, підтоплення, сніжні або безсніжні зими тощо). Серед екологів немає повної згоди в питанні про причини циклічних коливань чисельності, оскільки ці коливання надто упорядковані, щоб їх можна було відносити на рахунок одних лише змін у середовищі. Найвірогіднішим є пояснення, згідно з яким цикли пов'язані із взаємодією хижак-жертва. Відповідно до однієї з таких гіпотез збільшення популяції зайця супроводжується збільшенням популяції рисі, яка врештірешт досягає такої щільності, що популяція зайця не може витримати тиску хижака і починає скорочуватися. Слідом за цим починається зниження чисельності рисі, зумовлене скороченням запасу корму.
Однак цю гіпотезу, вважає американський еколог Р.Ріклефс, не можна прийняти як адекватне пояснення циклічних коливань популяцій рисі і зайця, тому що існує дуже багато даних, які їй суперечать. По-перше, репродуктивний потенціал у зайця значно вищий, ніж у рисі, а тому популяція рисі не може збільшуватися з такою швидкістю, яка б призвела до різкого скорочення популяції зайця, якщо якийсь інший фактор (наприклад, нестача корму) не зумовить зниження швидкості росту популяції зайця. По-друге, піки чисельності популяції рисі інколи збігаються з піками чисельності зайця або ж передують їм, а не слідують за ними з інтервалом один або декілька років. По-третє, на деяких острівцях, де немає рисі, спостерігаються такі ж коливання чисельності зайця-біляка, як і на материку. Можливо, що ці цикли чисельності популяції цих тварин зумовлені періодичними зниженнями якості і кількості рослин, якими живиться заєць, що, в свою чергу, призводить до скорочення популяції зайця (і популяції рисі), створюючи можливість для відновлення рослинності після надмірного виїдання її зайцями.
Дослідження динаміки чисельності лемінгів на Алясці підтвердили важливу роль песців у її пригніченні, але основна роль, як виявилось, належить якості корму. В рік, коли чисельність лемінгів досягала максимуму, вміст білка у рослинах, якими вони живляться, становив 22%, а в рік мінімуму - всього 14%
Стабільний тип охоплює популяції, які розвиваються в сталих умовах середовища і мають налагоджений механізм регулювання чисельності.
Вважають, що відносна стабільність в угрупованнях є наслідком функції популяцій окремих видів і невеликих груп контактуючих видів, а не видового багатства усього угруповання. Незмінність оточуючого середовища може забезпечити виживання багатьох видів, які впливають один на одного в умовах складного угруповання.
65
Американський еколог Р.Уіттекер на прикладі червоноокого віреона - основного виду дрібних птахів листопадних лісів на сході США показує, як можна визначити і виміряти "стабільність". На цій підставі, підкреслює автор, можна сформулювати деяку ідеальну концепцію і оцінити її в реальній ситуації. Допустимо, що віреон має усереднену щільність популяції, яка дорівнює 2000 особин на 1 км2 лісу, з яких 120 особин об'єднуються в 60 гніздуючих пар. Якщо смертність серед птахів становить 50% або 0,5 на рік, то кожний рік з 200 особин гине половина, або 100 птахів (під час міграції, стають здобиччю хижаків і т.п.). Щоб популяція залишилася стабільною, із 200 відкладених протягом літа яєць повинно вижити 100 пташенят, тобто 50%. Таким чином, популяція нагадує своєрідний "фонд", до якого особини прибувають і вибувають. Якщо швидкість надходження і загибелі особин однакова,-— підкреслює Уіттекер, - то загальну кількість птахів у популяції можна вважати стабільною, а сам цей стан стійким. В ідеалі "стабільність" популяції передбачає стійкий стан зі збалансованим у середньому рівнем народжуваності і смертності.
Які ж механізми регулювання відносної стабільності популяцій? Передусім це дотримання визначеної щільності популяції шляхом певних впливів: 1) знищують більшу частку особин (або зменшують народжуваність у розрахунку на кожну особину при рості популяції), і популяція таким чином досягає верхньої межі; 2) знищують меншу частку особин (або збільшують народжуваність на кожну особину) в умовах спаду щільності і таким чином встановлюють нижню межу її коливання. Для багатьох популяцій характерні спеціальні буферні механізми (такі, наприклад, як стадії спокою), які знижують витрати популяцій в періоди, коли абіотичне і біотичне середовища стають несприятливими для нормального розвитку.
Отже, робить висновок Уіттекер, не існує якогось одного механізму досягнення і підтримки відносної стабільності популяції. Деякі види є стабільними поряд з верхньою межею чисельності своєї популяції під впливом одного чи багатьох механізмів стійкості. Багато інших видів флуктує в широких межах, але продовжують існувати, оскільки ефект залежності від щільності або механізми буферності, які діють біля нижньої межі чисельності популяції, попереджують їх вимирання. Така відносна стабільність популяцій у природних угрупованнях є наслідком формування угрупованням протягом еволюції механізмів, які обмежують їх флуктуації.
66
2.4.2 Народжуваність і смертність. Тривалість життя
Народжуваність – це здатність популяції до збільшення. Термін "народжуваність" вживають і щодо людської популяції (в демографії). Він характеризує частоту появи нових особин будь-якого організму, незалежно від того, яким шляхом він з'являється: вилуплюється, відбруньковується, у процесі поділу тощо. Розрізняють максимальну і екологічну, чи реалізовану, народжуваність.
Плодючість характеризується коефіцієнтом народжуваності, тобто кількістю нащадків, які продукуються одиницею популяції за одиницю часу. Для людини він дорівнює приблизно 3,64%, тобто народжується 3,64 дитини на 100 мешканців на рік.
Смертність характеризує загибель особин у популяції. Аналогічно народжуваності смертність можна виразити кількістю особин, які загинули за даний період (кількість смертей за даний період). Екологічна, або реалізована смертність, загибель особин у даних умовах середовища,
– величина, яка, подібно до народжуваності, не є постійною, а змінюється залежно від умов середовища і стану самої популяції.
Виділяють значною мірою ідеальну величину – мінімальну смертність. Це постійна величина, що характеризує загибель особин в ідеальних умовах, при яких популяція не зазнає лімітуючих впливів. Максимальна тривалість життя особин у цих оптимальних умовах дорівнює її фізіологічній тривалості, яка в середньому значно перевищує екологічну тривалість життя. Часто значний інтерес являє собою не смертність, а виживання. Якщо кількість загиблих особин становить М, то виживання дорівнює 1 – М.
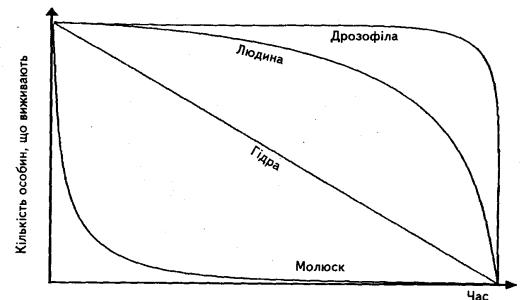
Від величини загибелі особин на різних фазах розвитку залежить загальний характер смертності популяцій, який може бути представлений відповідними типами кривих (рис.2.5)
Перший тип (виражений експоненціальною кривою) характеризується однаковими нормами смертності у кожному віці. Такий тип притаманний тваринам, які живуть у сталих оптимальних умовах, а також сучасній людині. Крива цього типу вирізняється дуже опуклою формою.
Другий тип відзначається підвищеною загибеллю молоді і відносною стійкістю дорослих особин. Такий характер смертності властивий більшості рослин і тварин, а також людині на ранніх стадіях розвитку.
Третій тип вирізняється стабільним існуванням молодих особин і різким підвищенням смертності дорослих, особливо старих особин.
Четвертий тип властивий метеликам, лососевим: здійснивши свої функції, вся генерація гине. У гідр смертність не змінюється протягом усього життя, а крива смертності наближається до прямої. Середня тривалість життя особин у популяції відповідає тривалості життя окремих
67
особин. Для життя популяції дуже важлива можливість розмноження її особин. Участь у цьому беруть особини репродукційного періоду. У тварин, як зазначено вище, розрізняють три стадії: передрепродукційну, або ювенільну, репродукційну і пострепродукційну, тобто період стерильності.
Рис.2.5 – Різні типи кривих виживання
Тривалість періодів у різних видів неоднакова. Часто періоду старості взагалі не буває, оскільки тварини гинуть у репродукційний період (комахи). Ювенільний період за тривалістю може дорівнювати репродукційному, а може бути значно довшим, ніж усе життя особини (стадія личинки у цикади триває 17 років, а доросла особина живе лише декілька тижнів).
Урослин виділяють чотири періоди розвитку особини: 1) латентний
–період первинного спокою – у вигляді насіння і плодів; 2) віргінільний - від проростання насіння до утворення генеративних органів; 3) генеративний – період розмноження насінням, спорами; 4) сенильний – період, коли рослини втрачають здатність до генеративного розмноження.
Окремі автори виділяють лише три фази: ювенільну, репродукційну і сенильну.
Оскільки народжуваність і смертність дуже змінюються з віком, відсоток різних вікових груп у популяції значною мірою визначає її репродуктивні можливості. Серед людей показник народжуваності є найвищим у віці близько 20 років. Найнижчий показник смертності у віці близько 12 років, а найвищий – протягом першого року життя і в старості. Ці характерні для кожного віку показники є сталими і, як правило, не змінюються, якщо незмінними залишаються умови життя. В історії людства двічі змінювалися рівні формування показників народжуваності і
68
смертності. Вперше народжуваність стала переважати смертність у період переходу від примітивного мисливського способу господарювання до рільництва, вдруге – в епоху загальної індустріалізації. Чисельність людей тривалий час утримувалася на низькому рівні і лише останні 500 років відбулося різке її збільшення (на початку нової ери – 230 млн, середньовіччя (V – середина XVII ст.) - 440 – 450 млн, 1800р. – 952млн, 1900р. – 1656 млн, 2000р. – близько 7 млрд).
Виділяють три типи вікових пірамід: з широкою основою і відповідно високим відсотком молодих особин, яка характерна для популяції з швидким ростом; з помірним відсотком молодих особин; з вузькою основою і чисельною перевагою старших особин над молодняком, що характерно для популяцій, які скорочуються.
Р. Ріклефс наводить приклад формування пірамід віку населення двох країн – Швеції і Коста-Ріки. Протягом багатьох десятиліть чисельність населення Швеції відносно постійна. Тут відзначається досить близька відповідність між віковим складом і очікуваною тривалістю життя. Кількість хворих людей відносно низька, що свідчить про незначне, але постійне переважання народжуваності над смертністю. В 1965 р. приріст населення становив 0,6%. Іншу картину спостерігаємо у КостаРіці, де приріст населення в 1963 р. становив 4,1%. Висока народжуваність різко збільшила молодші вікові класи, внаслідок чого і отримана піраміда з широкою основою.
Утварин з невеликою тривалістю життя приуроченість розмноження і смертності до певних сезонів створює циклічні зміни вікового складу. Наприклад, популяції медоносної бджоли навесні швидко збільшуються, і графік їх вікового складу набуває вигляду піраміди з широкою основою, характерною для популяцій, які ростуть. До кінця літа, в міру того як народжуваність сповільнюється, чисельність молодших вікових класів скорочується, а після припинення розмноження вони зникають зовсім. Бджоли, які пережили зиму, переходять в класи старшого віку.
Убагатьох тварин, які є об'єктом мисливства або рибальства, піраміди віку змінюються з волі людини. Тому дуже важливо професійно вести промисел популяцій, який можуть забезпечити лише досвідчені екологи.
Деколи гармонія між віковими групами відсутня, оскільки менша група не обов'язково кількісно переважає сусідню вищу групу. Наприклад, букові праліси Закарпаття з трьома-чотирма переважаючими віковими групами, які свідчать, що склад сучасної популяції визначається декількома хвилями масового розмноження дерев, розподілених у часі.
Важливим засобом аналізу вікового стану популяції є демографічні таблиці, які являють собою сукупність статистичної інформації про популяцію: кількість особин, яка доживає до кожного віку, і плодючість кожного вікового класу.
69
За допомогою демографічних таблиць вираховується швидкість росту популяції збільшення чисельності популяції відбувається шляхом множення (геометричний ріст), а не простого додавання (арифметичний ріст), тому що новонароджені молоді особини, виростаючи, самі продукують потомство. Такий ріст, за образним висловлюванням Р.Ріклефса, нагадує строковий вклад в ощадну касу, де наростаючі відсотки (новонароджене потомство) періодично додають до основного капіталу (дорослі особини, що розмножуються).
Як бачимо, народжуваність і смертність – це процеси, які відбивають взаємодію особини з оточуючим її середовищем, здатність популяції спрямувати наявні ресурси на розмноження, а також її здатність уникати хижаків, голоду або впливу несприятливих факторів.
Еколог, коли йдеться про демографію популяції, має виконати головне завдання: описати, витлумачити і зрозуміти закономірності поширення і динаміки чисельності живих істот. У зв'язку з цим необхідно підкреслити один безперечний факт, який можна записати у вигляді:
Nn = Nt + B – D + C – E. |
(2.6) |
Іншими словами, кількість особин певного виду, які заселяють дане місцезростання в даний час (Nn), дорівнює кількості особин, які проживали там раніше (N t), плюс кількість особин, які народилися протягом минулого відрізку часу (В), мінус кількість загиблих за той час особин (D), плюс кількість іммігрантів (С), мінус кількість емігрантів (Е).
Для еколога важливо з'ясувати причини, які впливають на величину Nn або ж передбачити величину N t. Вивчають стан рослинного і тваринного світу забрудненої річки, виміряють і реєструють N n цілого ряду видів, а потім порівнюють ці дані з тими, що одержані в незабрудненій річці. В кожному з цих випадків еколог досліджує стан демографічних процесів (народжуваність, смертність, міграції), які визначають чисельність, а також механізми впливу зовнішніх факторів на ці процеси.
2.4.3 Поліморфізм
Поліморфізм – існування в межах одного виду рослин або тварин двох (диморфізм) або більше груп особин з різко відмінними ознаками. Розрізняють сезонний, статевий і віковий поліморфізми.
Сезонний поліморфізм – відмінність особин різних поколінь, що розвиваються в різні пори року (літня і зимова форми дуба звичайного, літнє і зимове забарвлення зайця-русака).
Статевий поліморфізм – у бджіл – робочі бджоли, матки, трутні; у термітів – матки, робітники, солдати.
70
Віковий поліморфізм – у комах – яйце-личинка, лялечка, доросла особина.
Розрізняють ще фенотипічний поліморфізм, пов'язаний з пристосуванням рослин до таких явищ, як зміна сезону.
Поліморфізм, як наслідок еволюції видів, має велике біологічне значення, оскільки сприяє існуванню виду в дуже відмінних умовах, а також відкриває шлях до утворення нових видів. У останні десятиліття вчені навчилися виявляти пов'язану із відбором мінливість всередині невеликих локальних популяцій. Така мінливість відома як генетичний поліморфізм – співіснування в межах одного і того ж місцезростання двох або більше виразно відмінних внутрівидових форм, причому в таких співвідношеннях, що постійна присутність найрідкісніших з цих форм не може бути віднесена лише на рахунок безперервного мутагенезу та міграцій.
Однак далеко не всі прояви такого характеру мінливості, на думку окремих вчених, відбивають певні відповідності між організмами і середовищем, навпаки, деякі з них виявляють елементарні неузгодженості. Вони виникають тоді, коли одна спеціалізована форма на стадії розселення проникає у місцезростання іншої, а також тоді, коли при зміні умов одна форма витісняється іншою, краще пристосованою до змінених умов місцезростання. Такий поліморфізм називають перехідним. Цей перехідний характер пояснюється тим, що жодна популяція не може встигнути за змінами умов місцезростання і водночас не може їм запобігти.
Однак у багатьох випадках внутріпопуляційний поліморфізм активно підтримується природним відбором різними способами, зокрема:
а) в деяких випадках гетерозиготи визначаються підвищеною пристосованістю, але внаслідок менделевого розщеплення вони постійно поповнюють популяцію породженими ними менш життєздатними гомозиготами. Цей "гетерозис" пояснює існування серпоподібноклітинної анемії в популяціях людини, яка трапляється у вогнищах малярії. Індивідууми, гетерозиготні стосовно локусу, страждають легким недокрів'ям, але рідко хворіють на малярію. При цьому вони безперервно породжують гомозиготних індивідуумів – або уражених тяжким недокрів'ям, або ж сприйнятливих до малярії;
б) інтенсивність відбору може змінюватися в межах деякого діапазону, причому на одній із його меж відбір може сприяти одній формі (морфі), а на іншій межі – іншій. При проміжній інтенсивності відбору можуть виникати поліморфні популяції.
в) деколи відбір буває частотно-залежним: будь-яка внутрівидова форма найжиттєздатніша тоді, коли вона трапляється найрідше. Вважають, що саме тому виживають незвично забарвлені
71
форми: вони життєздатні, оскільки хижаки їх не розпізнають і не чіпають;
г) дрібномасштабна просторова структура популяції і її місцезростання бувають дуже складними, і в різних частинках цієї "латаної ковдри" (мозаїки) відбір може відбутися в різних напрямах. Збереження відповідності між організмом і середовищем у такій ситуації неминуче залежить від розсіювання численних розселювальних стадій: якщо чисельність їх достатньо велика, то достатньо велика і вірогідність того, що частина їх укорінюється саме в цьому "клапті", де відповідна форма найстійкіша. Можливий і інший варіант: організм може бути довговічним і здатним переміщатися, використовуючи при цьому умови і ресурси найпридатніших "клаптів".
Англійські вчені Р.Туркінгтон і Дж.Харпер вивчали, якою мірою властивості різновидів конюшини повзучої (Тrifolium repens) відбивають локалізовані відповідності між рослинами і локальними особливостями місцезростань. Вони викопували рослини з поля, розмножували їх у теплиці, а потім знову висаджували в поле. Одну частину кожного з клонів вони повертали туди, звідки було взято родинну рослину, а інші частини туди, звідки походили інші клони. Спочатку вибірки були взяті з ділянок, на яких переважали різні лучні трави. Потомство кожної рослини висаджували на ділянки з різнотрав'ям. Після пересадки усі різновиди конюшини краще за все росли на полі там, де вони межували з тими ж травами, що й до пересадки в теплицю. Ці результати прямо і дуже переконливо свідчать на користь того, що різні генотипні різновиди конюшини розподілені по пасовищу таким чином, що кожна з них відповідає локальним умовам існування.
На тому ж полі Р.Дірзо і Дж.Харпер (1982) досліджували розподіл форм конюшин, які відбивають наявність ще одного специфічного типу поліморфізму, який полягає у тому, що при пошкодженні (наприклад, при скошуванні) одні рослини виділяють синильну кислоту, а інші не виділяють. Відомо, що слимаки звичайно не поїдають конюшину, що містить ціаніди. Між розподілом ціановмісної форми конюшини і ділянок з підвищеною щільністю слимаків був виявлений високодостовірний зв'язок.
Встановлена ще одна внутріпопуляційна відміна за розселеністю (фенотипічна) – поліморфізм потомства одного і того ж родича, котрий є наслідком неоднорідності і мінливості умов місцезростання, в яких, наприклад, рослини намагаються вижити, залишаючись на межі (чекаючи сприятливого року чи сезону) або ж "кочуючи" в пошуках цих же сприятливих умов. Зразків такого диморфізму чимало серед квіткових рослин, особливо в родинах злакових, складноцвітих, лободових і хрестоцвітих. У всіх цих випадках причиною відмін насіння є не генетичне походження, а батьківські рослини. І з того насіння, що
72