

Биохимия растений
.pdfной жирной кислоты с одной двойной связью выход АТФ умень˝- шается на два моля, с двумя — на четыре, с тремя двойными св˝язями — на шесть молей. Следовательно, в процессе окисления ˝в митохондриях 1 моля линоленовой кислоты, содержащей три
двойные связи, выход АТФ по сравнению со стеариновой кисл˝о- той будет составлять не 147 молей, а 141. В расчете на 1 г окисляе˝-
мой линоленовой кислоты выход АТФ составит 0,51 моля, т. е. значительно больше, чем для глюкозы и других углеводов.
Как было показано ранее, определенная часть жирных кис-
лот, образующихся при распаде жиров, у растений подвергае˝тся
α-окислению в цитоплазме и при каждом обороте α-окисления
происходит отщепление от жирной кислоты одной молекулы С˝О
2
и синтез одной молекулы восстановленного динуклеотида НАД · Н.
Следовательно, если даже предположить, что происходит пол˝ное
окисление жирной кислоты, то может осуществиться синтез т˝ако-
го числа молекул НАД · Н, которое на один меньше числа угле-
родных атомов в молекуле жирной кислоты.
Например, для молекулы стеариновой кислоты оно будет рав-˝
но 17. Если сделать пересчет синтезированных молекул НАД · Н в
эквиваленты АТФ, принятые для системы окислительного фос˝фо-
рилирования, то мы получим выход АТФ значительно меньший,˝
чем при окислении жирных кислот по механизму β-окисления и последующего окисления ацетил-КоА в цикле Кребса. Однако
надо иметь в виду, что бó льшая часть восстановленных динуклео-
тидов НАД · Н, образуемых в цитоплазме по механизму α-окисле-
ния, в системе окислительного фосфорилирования митохонд˝рий
не окисляется, а используется в восстановительных реакци˝ях, происходящих в цитоплазме.
10.5.ПРЕВРАЩЕНИЕ ЖИРНЫХ КИСЛОТ
ÂУГЛЕВОДЫ
Óрастений процесс распада жиров интенсивно происходит п˝ри прорастании семян, в которых они служат основным запасным˝ ве-
ществом. При этом часть запасного жира в прорастающих сем˝енах
расходуется как энергетический материал для инициации с˝опряженного синтеза органических веществ, необходимых для жи˝знедеятельности развивающихся проростков, а другая часть яв˝ляется источником органических метаболитов для построения мол˝екул структурных и функционально активных компонентов их кле˝ток.
Важнейший путь использования жиров в качестве биохимиче˝ских
метаболитов — это превращение их в углеводы, которое осу˝ществ-
ляется через реакции глиоксилатного цикла, называемого также
циклом глиоксиловой кислоты.
411
Ферменты, катализирующие реакции глиоксилатного цикла, локализованы в специализированных внутриклеточных стру˝ктурах — глиоксисомах, которые функционально связаны с митохондриями. Участие глиоксисом в превращении жирных кислот в у˝гле-
воды наиболее хорошо изучено при прорастании семян масли˝ч- ных растений. Глиоксисомы формируются в начале прорастан˝ия
этих семян и деградируют после полного расщепления жира. Кроме ферментов глиоксилатного цикла в глиоксисомах так˝же
содержатся ферменты, катализирующие β-окисление жирных
кислот. Продукт β-окисления жирных кислот — ацетил-КоА — да-
лее включается в реакции глиоксилатного цикла.
Первые три реакции этого цикла такие же, как и в цикле Креб-˝ са. Вначале ацетил-КоА реагирует с енольной формой щевелево-
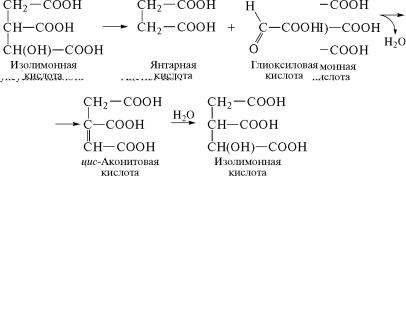
уксусной кислоты, образуя лимонную кислоту. Затем последо˝ва-
тельно происходит превращение лимонной кислоты в öèñ-акони-
товую и изолимонную кислоты:
На следующем этапе превращений изолимонная кислота расщепляется ферментом изоцитратлиазой (4.1.3.1) на два продук-
та — янтарную и глиоксиловую кислоты:
Изоцитратлиаза — аллостерический фермент, активность к˝ото-
рого подавляется избыточной концентрацией фосфоенолпир˝ови-
ноградной кислоты, образующейся в цитоплазме из продукто˝в превращения янтарной кислоты. Последняя для дальнейших р˝е-
412

акций поступает в митохондрию. Глиоксиловая же кислота в ˝глиоксисоме вступает во взаимодействие с новой молекулой ац˝етилКоА, образуя яблочную кислоту. Эту реакцию катализирует фермент малатсинтаза (4.1.3.2):
В последней реакции глиоксилатного цикла яблочная кисло˝та
окисляется ферментом малатдегидрогеназой в щавелевоуксусную кислоту:
После этого щавелевоуксусная кислота изомеризуется в свою
енольную форму и может снова вступать в первую реакцию ци˝кла. Как видно из представленных ранее реакций, в глиоксилатны˝й
цикл кроме регенерирующегося субстрата — енольной форм˝ы ща-
велевоуксусной кислоты — включаются две молекулы ацети˝л-
КоА, а продуктами цикла являются янтарная кислота и НАД ·˝ Н.
Поэтому суммарное уравнение глиоксилатного цикла можно˝ за-
писать в следующем виде:
Янтарная кислота, поступающая из глиоксисомы в митохондрию, включается там в реакции цикла Кребса и превращается˝ в
щавелевоуксусную кислоту, которая далее из митохондрии т˝ранс-
портируется в цитоплазму, где c участием соответствующих ˝ферментов используется для синтеза углеводов.
Под действием фермента фосфоенолпируваткарбоксикиназы
(4.1.1.32) щавелевоуксусная кислота с участием ГТФ подвергаетс˝я декарбоксилированию, превращаясь в фосфоенолпировиногр˝ад-
ную кислоту:
413

Образовавшаяся фосфоенолпировиноградная кислота далее˝
включается в уже известный нам цикл реакций обращения гли˝колиза (см. с. 327—330), конечным продуктом которого является фруктозо-6-фосфат. Из фруктозо-6-фосфата могут синтезироват˝ь-
ся различные углеводы. Но если фруктозо-6-фосфат вступает в˝ дыхательные реакции, дающие кетокислоты, то возможно его ˝использование и для синтеза аминокислот. Общая схема превра˝щения жиров в углеводы показана на рисунке 10.1.
У бактерий реакции глиоксилатного цикла пространственн˝о не отделены от цикла Кребса, поэтому глиоксилатный путь прев˝ращений ацетил-КоА можно рассматривать как видоизмененный˝
цикл трикарбоновых кислот. У человека и животных глиоксил˝атный цикл отсутствует, поэтому у данных организмов отложен˝ный в жировых тканях жир не может так легко использоваться дл˝я превращения в углеводы, как у бактерий или растений.
Весь процесс биосинтеза углеводов из продуктов распада ж˝ира
получил название глюконеогенеза. Такое название он имеет вследствие того, что в ходе обращения реакций гликолиза из прод˝укта распада жирных кислот фосфоенолпировиноградной кислоты˝
осуществляется синтез важнейшего представителя сахаров˝ — глюкозы, из которой уже довольно легко могут синтезироваться˝ дру-
гие углеводы.
В глиоксисоме |
В цитоïлаçме |
|||
Жирные кислоты |
Äрóгие óглеâоды |
|||
|
β-окисление |
|
|
|
|
|
|
|
|
Ацетил-КоА |
Ôрóктоçо-6- |
ôосôат |
||
|
|
|
|
|
Глиоксилатный цикл |
Òриоçоôосôаты |
|||
|
|
|
||
|
|
|
|
|
Янтарная кислота |
Ôосôоенолïироâино- |
|||
|
|
градная кислота |
||
|
|
|||
|
|
|
ÑO |
|
|
|
ГÒÔ |
2 |
|
Цикл Кребса |
Ùаâелеâо- - |
|||
В митохондрии |
óксóсная |
|
||
кислота |
|
|||
|
|
|
||
Рис. 10.1. Превращение жирных кислот в углеводы с участием гл˝иоксисом и митохондрий
414
10.6. РАСПАД ФОСФОЛИПИДОВ
Áó льшая часть фосфолипидов находится в клетках листьев в
связанном состоянии, так как они участвуют в построении к˝ле-
точных мембран. Переход их в свободное состояние может пр˝оис-
ходить при перестройке и обновлении этих структур. В несв˝язан-
ном (свободном) состоянии много фосфолипидов откладывае˝тся в
семенах, где они используются в качестве запасных веществ˝. При
прорастании семян наблюдается высокая активность гидро˝лити- ческих ферментов, катализирующих распад запасных вещест˝в, в
том числе и запасных фосфолипидов, главным образом фосфат˝и-
дилэтаноламинов и фосфатидилхолинов.
Распад фосфолипидов начинается с действия фосфолипаз, êàòà-
лизирующих гидролиз сложноэфирных связей глицерина и жи˝р- ных кислот. Вначале происходит отщепление остатка жирной˝
кислоты, связанного со вторым углеродным атомом, а затем с˝вя-
занного с первым углеродным атомом. В результате действия˝ этих
ферментов образуются свободные жирные кислоты и соедине˝ния
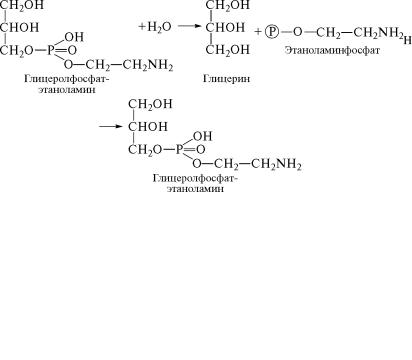
азотистого основания с глицеролфосфатом:
На следующем этапе под действием специфической фосфатаз˝ы гидролизу подвергается сложноэфирная связь между остат˝ком глицерина и фосфатом азотистого основания:
415
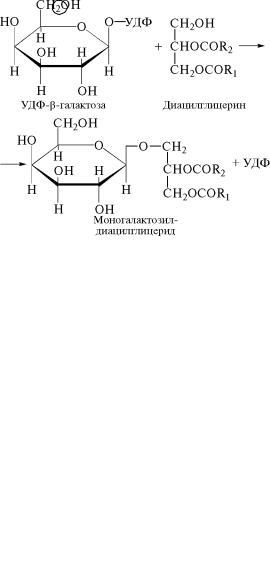
Гидролиз образовавшегося этаноламинфосфата также катал˝и- зирует специфическая фосфатаза:
P—O—CH2 —CH2NH2 +
+H2O ¾¾¾¾Фосфатаза¾® ÍΗÑH2 —CH2 NH2 +H3PO4
Этаноламин
Под действием указанных ферментов осуществляется превр˝а-
щение фосфолипидов в свободный глицерин, жирные кислоты, азотистые основания и неорганический фосфат, которые дал˝ее
могут включаться в различные биохимические реакции или о˝кис-
ляться в процессе дыхания.
10.7. СИНТЕЗ И ПРЕВРАЩЕНИЯ ДРУГИХ ЛИПИДОВ
Синтезируемые в хлоропластах растений галактолипиды об˝ра-
зуются в ходе реакций переноса остатков галактозы от УДФ-˝га-
лактозы на диацилглицерины. Если осуществляется перенос˝ одного остатка галактозы в виде b-пиранозы на диацилглицерин,
синтезируются молекулы b-галактозилдиацилглицеридов:
416
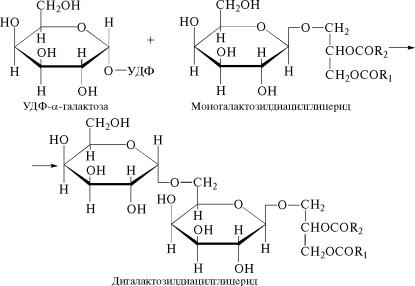
При переносе на моногалактозилдиацилглицерид еще одног˝о остатка галактозы в виде α-пиранозы осуществляется синтез дигалактозилдиацилглицеридов:
Следует отметить, что в процессе синтеза диацилглицерино˝в,
входящих в состав гликолипидов, к первому углеродному ато˝му
глицерина присоединяется остаток насыщенной жирной кис˝лоты
(содержащей радикал R1), а ко второму углеродному атому пре-
имущественно остаток линоленовой кислоты (содержащей ра˝ди-
êàë R2).
При перестройке и обновлении структуры хлоропластных ме˝м- бран образуются свободные формы гликолипидов, которые мо˝гут
подвергаться превращениям в результате действия соотве˝тствую-
щих гидролаз. Отщепление остатков моносахаридов катализ˝ируют специфические гликозидазы, а остатков жирных кислот — г˝идролазы сложных эфиров. Механизмы этих реакций в растительны˝х клетках пока еще недостаточно изучены.
Исходным соединением для синтеза стеролов служит продук˝т обмена углеводов и жирных кислот ацетилкофермент А, включ˝а-
ясь в обмен терпеноидных соединений он образует сквален (см. с. 564), из которого через ряд промежуточных стадий синтезир˝у-
ются стеролы. Важнейшим промежуточным продуктом в этих ре˝-
акциях у растений является циклоартенол, который служит важ-
417
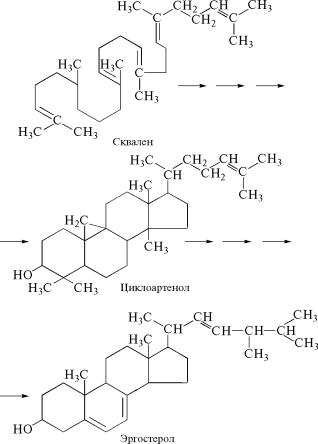
нейшим биохимическим предшественником при образовании ˝стеролов из сквалена.
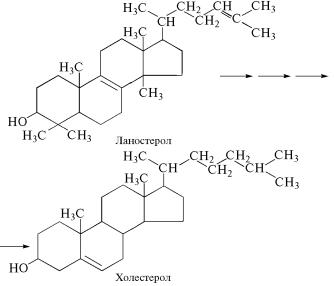
В организмах животных стероидные липиды синтезируются и˝з сквалена, однако непосредственным биохимическим предше˝- ственником для их образования служит ланостерол, который через
ряд промежуточных стадий превращается в холестерол. Другие стероидные вещества животного организма образуются пут˝ем пре-
вращений холестерола.
418
Распад стероидных липидов — сложный многоступенчатый
процесс, который пока еще слабо изучен. Однако известно, чт˝о
конечным продуктом распада стеролов, как и при распаде др˝у- гих липидов, является ацетилкофермент А. Еще меньше сведе-
ний имеется по обмену липидов воска. В ряде опытов было по-
казано, что главные липидные компоненты воска — высокомолекулярные одноатомные спирты образуются из пальмитино˝вой
кислоты.
Контрольные вопросы. 1. Из каких основных компонентов и с участием каких ферментов синтезируются ацилглицерины жиров? 2. Как проис˝ходит синтез и распад глицерина? 3. Каковы биохимические механизмы синте˝за насыщенных жирных кислот? 4. Каковы особенности образования жирных ки˝слот с четным и нечетным числом углеродных атомов, а также с боковыми отв˝етвлениями углеродной цепи? 5. Как осуществляется превращение насыщенных˝ кислот в ненасыщенные? 6. С помощью каких показателей можно оценивать ин˝тенсивность синтеза жиров и ненасыщенных жирных кислот в семенах масл˝ичных растений? 7. Какие биохимические реакции лежат в основе синтеза фосфо- и гликолипидов? 8. С участием каких ферментов осуществляется распад ац˝илглицеринов и фосфолипидов? 9. Каков механизм α-окисления жирных кислот? 10. Какова последовательность биохимических реакций β-окисления насыщенных и ненасыщенных жирных кислот? 11. Каковы особенности биодеградац˝ии жирных кислот с нечетным числом углеродных атомов и боковыми отв˝етвлениями в углеродной цепи? 12. К каким экологическим нарушениям приводи˝т использова-
419
ние в сельском хозяйстве препаратов, содержащих жирные ки˝слоты с боковыми ответвлениями у нечетных углеродных атомов? 13. Какие биоэн˝ергетические продукты образуются в ходе α- è β-окисления жирных кислот и каков суммарный энергетический эффект окисления жирных кислот? 14. Как п˝роисходит в растениях превращение жирных кислот в углеводы? 15. Какова п˝оследовательность реакций глиоксилатного цикла и в каких клеточных ст˝руктурах проходят эти реакции? 16. По каким механизмам из продукта глиоксилатного цикла янтарной кислоты синтезируются углеводы? 17. Как осуществляются синтез и превращения стероидных липидов? 18. Что известно о природе обра˝зования липидных компонентов воска?
420