

Биохимия растений
.pdfпоступать в любые акцепторные клетки растительного орга˝низма и использоваться в них для синтеза различных органически˝х веществ.
Таким образом, у растений наряду с процессом фотосинтеза
существует дополнительный механизм углеродного питания˝ за счет СО2 почвы, поступающего в нефотосинтезирующие клетки
корней. Однако первичный акцептор СО2 для этих реакций — фосфоенолпировиноградная кислота образуется в результа˝те
окисления фотоассимилятов, поступающих в корни из листье˝в.
9.3. ДЫХАНИЕ
Основной источник энергии для поддержания процессов жиз˝-
недеятельности у гетеротрофных организмов — это химиче˝ская энергия органических веществ, которые они используют в ка˝че-
стве пищи. Несинтезирующие органы растений, а в темноте и
способные к фотосинтезу клетки, в качестве источника энер˝гии
используют также органические вещества, образующиеся ка˝к про-
дукты фотосинтеза. Для того чтобы химическая энергия орга˝ни- ческих веществ могла использоваться для поддержания жиз˝недея-
тельности организма, они подвергаются окислению. При этом˝ на
каждом этапе окислительного распада веществ выделяется˝ энер-
гия, которая связывается в ходе определенных реакций в ви˝де
макроэргических соединений и восстановленных динуклеот˝идов и может затем использоваться для осуществления эндергон˝ичес-
ких процессов, связанных с синтезом, транспортом и активи˝рова-
нием различных химических веществ. В ходе биологического˝
окисления образуются промежуточные метаболиты, которые˝ затем участвуют в синтезе конечных продуктов, выполняющих в организме те или иные биологические функции (структурные˝, запасные вещества и др.).
Последовательность биохимических реакций, лежащих в осн˝о-
ве биологического окисления, называют дыханием. У растений и других высших организмов в реакциях дыхания участвует ки˝слород, а в качестве продуктов дыхания выделяются СО2 è Í2О. Первые сведения о дыхании растений были получены Я. Ингенхау˝зом
(1778), а к концу XIX в. было установлено общее уравнение дыха-
тельного процесса:
Ñ6H12O6 + 6O2 → 6CO2 + 6H2O + G, |
(1) |
ãäå Ñ6Í12Î6 — моносахариды, образуемые при фотосинтезе.
В дальнейшем благодаря работам В. И. Палладина (1912) было
показано, что на первом этапе дыхательных реакций происхо˝дит
321

отщепление водорода от молекул сахара и воды и образовани˝е промежуточных восстановленных продуктов, а на следующем˝ этапе эти продукты уже окисляются кислородом с образованием˝ воды. Поэтому дыхательный процесс можно представить в две
стадии:
Ñ6H12O6 + 6Í2Î → 6CO2 + 24[H] |
(2) |
24[H] + 6O2 → 12H2O |
(3) |
Если из уравнений (2) и (3) составить суммарное уравнение процесса, то мы получим уравнение (1).
В 1921—1925 гг. в опытах по изучению роли фермента цитохромоксидазы О. Г. Варбург и Д. Кейлин установили, что на второй ста-
дии дыхания на кислород переносятся электроны и протоны, ˝ко-
торые и участвуют в образовании молекул воды:
Î2 +4å +4Í+ → 2Í2Î
Отщепление электронов и протонов от молекул сахара и воды˝,
как было выявлено в ходе дальнейших исследований, катализ˝иру-
ют ферменты дегидрогеназы, и с их участием происходит образо-
вание восстановленных динуклеотидов НАД · Н и ФАД · Н2, при окислении которых и осуществляется перенос электронов и˝ про-
тонов на кислород.
По современным представлениям, процесс дыхания у растени˝й
состоит из трех стадий: гликолиза, цикла ди- и трикарбоновы˝х
кислот, а также окисления восстановленных динуклеотидов˝, образующихся на первых двух стадиях дыхания, с участием ферме˝нт-
ных комплексов, локализованных во внутренних мембранах м˝ито-
хондрий.
9.3.1. ГЛИКОЛИЗ
Гликолиз представляет собой анаэробную стадию дыхания у
высших организмов и основной механизм дыхания у анаэробн˝ых
микроорганизмов. В процесс гликолиза включаются активированные формы моносахаридов — гексоз, которые подвергаются˝ окислительному распаду с образованием пировиноградной кисл˝оты. Большой вклад в раскрытие механизмов этих реакций внесли˝ из-
вестные биохимики Г. Эмбден, О. Ф. Мейергоф и Я. О. Парнас. Реакции гликолиза локализованы в жидкой фазе цитоплазмы˝ и
хлоропластов. Ферменты, катализирующие эти реакции в цито˝-
плазме, образуют комплексы с актиновыми белками цитоплаз˝ма-
тических филаментов, таким образом обеспечивается локал˝изация происходящих превращений.
322

Активирование гексоз осуществляется путем фосфорилиров˝а- ния с участием АТФ под действием соответствующих киназ, и об-
разующиеся фосфаты гексоз изомеризируются во фруктозо-6-
фосфат, который уже далее включается в реакции гликолиза.˝ В первой реакции фруктозо-6-фосфат подвергается повторному˝ фосфорилированию под действием фермента фосфофруктокиназы:
(1)
Образующийся в этой реакции фруктозо-1,6-дифосфат имеет
полярную молекулу, которая легко подвергается расщеплен˝ию на
следующем этапе превращений.
Фосфофруктокиназа — аллостерический фермент и поэтому˝
подвержен активной регуляции. Его активность ингибирует˝ся вы-
сокой концентрацией АТФ и лимонной кислоты, образующейся˝ в
цикле Кребса. Однако фермент аллостерически активируетс˝я
АДФ. Следовательно, при высокой обеспеченности клетки эне˝р- гией, повышающей концентрацию АТФ, происходит замедление˝
первой реакции гликолиза и в целом интенсивности дыхания˝.
Когда же энергетический потенциал клетки понижается и во˝зрас-
тает концентрация АДФ, последний аллостерически активир˝ует
фермент фосфофруктокиназу, вследствие чего возрастает с˝корость
включения фруктозо-6-фосфата в реакции гликолиза, вызывая˝
повышение интенсивности дыхания, обеспечивающего клетк˝у
энергией.
На следующем этапе гликолиза под действием фермента альдолазы происходит расщепление фруктозо-1,6-дифосфата на две триозы — 3-фосфоглицериновый альдегид и фосфодиоксиаце˝-
òîí:
(2)
323

Между триозофосфатами устанавливается динамическое рав˝- новесие с участием фермента триозофосфатизомеразы:
(3)
Образовавшийся 3-фосфоглицериновый альдегид далее окисляется в 3-фосфоглицериновую кислоту под действием фермента
глицеральдегидфосфатдегидрогеназы (1.2.1.12), молекулы которого
состоят из четырех полипептидных субъединиц с молекуляр˝ными
массами по 45 000 и содержат в активном центре тиоловые групп˝ы
(—SН) и окисленные динуклеотидные группировки НАД+. При взаимодействии фермента с 3-фосфоглицериновым альдегидо˝м
образуется фермент-субстратный комплекс, в котором иници˝иру-
ется возникновение ацилмеркаптановой макроэргической с˝вязи и
восстановление динуклеотидной группировки НАД+ â ÍÀÄ · Í:
(4)
После этого фермент-субстратный комплекс подвергается ф˝ос-
форолизу с участием неорганического ортофосфата. При это˝м НS-
фермент отщепляется от субстрата и к остатку последнего п˝рисоединяется фосфатная группировка, в которой за счет энергии˝ ацилмеркаптановой связи возникает макроэргическая ацилфосф˝атная связь:
(5)
324

В следующей реакции остаток фосфорной кислоты с макроэргической связью переносится на АДФ, в результате чего осу˝ществляется синтез АТФ. Эту реакцию катализирует фермент фосфоглицераткиназа (2.7.2.3):
(6)
Так как в реакции (5) макроэргическая ацилфосфатная связь
возникает в составе субстрата, такой процесс называют субстратным фосфорилированием.
Полученная в реакции (6) 3-фосфоглицериновая кислота под действием фермента фосфоглицератмутазы изомеризуется затем в
2-фосфоглицериновую кислоту:
(7)
После этого от 2-фосфоглицериновой кислоты отщепляется молекула воды с образованием фосфоенолпировиноградной ˝кислоты. Реакцию дегидратации катализирует фермент енолаза (4.2.1.11):
(8)
325

В результате дегидратации в молекуле субстрата происход˝ит перераспределение внутренней энергии и возникает макро˝эрги- ческая связь.
От молекулы фосфоенолпировиноградной кислоты фосфатная˝
группировка с макроэргической связью передается на АДФ, в˝ ре-
зультате чего осуществляется синтез АТФ. Перенос фосфатн˝ой
группировки в этой реакции катализирует фермент пируваткиназа
(2.7.1.40):
(9)
Пируваткиназа аллостерически ингибируется высокой конц˝ен-
трацией АТФ и ацетилкофермента А, который образуется при
дальнейшем превращении пировиноградной кислоты в ходе д˝ыха-
тельных реакций.
Продукт реакции (9) енолпировиноградная кислота самопроизвольно подвергается перегруппировке и превращается в˝ более
устойчивую кетоформу — пировиноградную кислоту:
(10)
Учитывая, что из гексозы образуется две триозы, в ходе их
окисления возникают две молекулы пировиноградной кисло˝ты.
По этой же причине все реакции гликолиза, начиная с четвер˝той, при окислении одной молекулы гексозы повторяются дважды˝. В ходе гликолиза, кроме двух молекул пировиноградной кисло˝ты, синтезируются биоэнергетические продукты в виде четыре˝х молекул АТФ [в реакциях (6) и (9)] и двух молекул НАД · Н [в реакции
(4)]. Поскольку на активирование гексозы затрачивается две˝ моле-
кулы АТФ, окончательный выход АТФ при гликолизе составляе˝т не четыре, а две молекулы. Кроме того, в реакции (9) происходи˝т
дегидратация 2-фосфоглицериновой кислоты, поэтому в расче˝те
326
на одну молекулу окисляемой гексозы выделяется две молек˝улы воды. Исходя из этих данных, суммарное уравнение гликолиз˝а принимает следующий вид:
C H O |
6 |
+2ÍÀÄ+ +2Í ÐÎ +2ÀÄÔ |
→ 2ÑH —CO—COOH + |
||
6 |
12 |
3 |
4 |
3 |
|
Гексоза |
|
|
|
Пировиноградная |
|
|
|
|
|
|
кислота |
+2ÍÀÄ·Í+2Í+ +2ÀÒÔ+2Í2Î
Конечный продукт гликолиза — пировиноградная кислота представляет собой один из важнейших продуктов дыхания и˝ в
целом обмена веществ любого организма. У анаэробных орган˝измов пировиноградная кислота включается в реакции брожен˝ия, в ходе которых в зависимости от организма происходит синте˝з эта-
нола, молочной, уксусной, пропионовой кислот и некоторых д˝ругих биохимических продуктов. У растений и других аэробных˝
организмов пировиноградная кислота подвергается окисле˝нию до СО2 è Í2О в ходе реакций цикла Кребса. В некоторых органах растений при недостатке кислорода также могут частично про˝исхо-
дить реакции брожения, конечным продуктом которых чаще вс˝его является этиловый спирт.
Через образование пировиноградной кислоты, как будет показано далее, осуществляется связь между обменом углевод˝ов, липидов и белков в растительном организме. Из пировиногра˝д-
ной кислоты путем аминирования синтезируется аминокисл˝ота аланин, которая участвует в реакциях азотного обмена и си˝нтезе
белков. Путем превращения в ацетил-КоА пировиноградная ки˝с-
лота дает начало синтезу жирных кислот, а при распаде белк˝ов, аминокислот, жирных кислот на определенных этапах образу˝ет-
ся пировиноградная кислота, которая может использоватьс˝я на
синтез углеводов путем обращения реакций гликолиза благ˝одаря тому, что большинство ферментов, катализирующих эти реак-˝
ции, обладают обратимым действием. Однако некоторые стади˝и
гликолиза необратимы и для их преодоления используются о˝б- ходные пути.
Первый необратимый этап в ходе превращения пировиноградной кислоты в гексозу — это образование из пировиног˝радной кислоты фосфоенолпировиноградной. Он осуществляетс˝я в четыре последовательные реакции, происходящие в митохон˝дриях и цитоплазме. Вначале пировиноградная кислота карбокс˝или-
руется с участием АТФ под действием митохондриальной пируваткарбоксилазы (6.4.1.1) и превращается в щавелевоуксусную
кислоту:
327

Затем щавелевоуксусная кислота восстанавливается митохондриальной малатдегидрогеназой в яблочную кислоту:
Образовавшаяся яблочная кислота перемещается в цитопла˝зму
и под действием цитоплазматической малатдегидрогеназы снова
превращается в щавелевоуксусную кислоту:
После этого под действием фермента фосфопируваткарбоксикиназы (4.1.1.32) из щавелевоуксусной кислоты путем декарбоксилирования образуется фосфоенолпировиноградная кислота:
Превращение фосфоенолпировиноградной кислоты в 3-фос-
фоглицериновый альдегид осуществляется в результате об˝раще-
ния соответствующих реакций гликолиза (8—4). В ходе этих
реакций на фосфорилирование 3-фосфоглицериновой кислоты
затрачивается АТФ, а на восстановление фермент-субстратн˝ого
комплекса глицеральдегидфосфатдегидрогеназы — восстан˝овленные динуклеотиды НАД · Н.
328
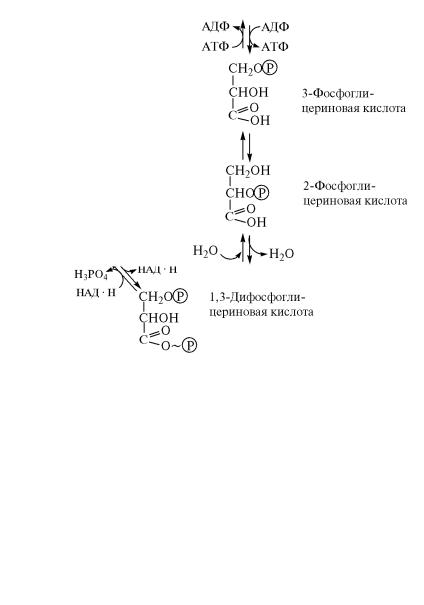
Рис. 9.7. Реакции гликолиза и обходные пути их обращения
329

Рис. 9.7. Реакции гликолиза и обходные пути их обращения. Про˝должение
В результате обращения реакций гликолиза (3) и (2) происхо-
дит синтез фруктозо-1,6-дифосфата из 3-фосфоглицеринового
альдегида, а под действием фруктозо-1,6-дифосфатазы фруктоз˝о-
дифосфат гидролизуется с образованием фруктозо-6-фосфата˝, с
превращения которого начинаются реакции гликолиза:
Реакции гликолитического окисления фруктозо-6-фосфата до˝ пировиноградной кислоты и обратный процесс синтеза фрук˝тозо-
6-фосфата из пировиноградной кислоты могут быть представл˝ены
в виде следующей схемы (рис. 9.7).
Реакции гликолиза являются для функционирующей клетки важным источником биоэнергетических продуктов, таких ка˝к
АТФ и НАД · Н, а также промежуточных метаболитов, которые˝ могут использоваться для синтеза многих жизненно важных˝ ве-
330