

Биохимия растений
.pdf9.1.3. ФОТОДЫХАНИЕ
Изучение механизма действия фермента рибулозодифосфат-˝
карбоксилазы показало, что конкурентным ингибитором это˝го фермента является кислород, который конкурирует с СО2 ïðè âçà-
имодействии последнего с каталитическим центром фермен˝тного
белка. Поэтому при высокой концентрации кислорода и низко˝й концентрации СО2 в воздухе карбоксилирующая активность рибу-
лозодифосфаткарбоксилазы понижается, но усиливается ее˝ окси-
геназная способность, вследствие чего к рибулозо-1,5-дифосф˝ату
присоединяется не СО2, а кислород, в результате происходит рас-
щепление рибулозо-1,5-дифосфата на 3-фосфоглицериновую и фосфогликолевую кислоты:
Образовавшаяся фосфогликолевая кислота под действием с˝пе-
цифической фосфатазы подвергается гидролизу с образова˝нием
неорганического фосфата и гликолевой кислоты:
Гликолевая кислота подвергается дальнейшим превращения˝м в пероксисомах — субклеточных органеллах, функциональная дея-
тельность которых тесно связана с процессами, происходящ˝ими в
хлоропластах и митохондриях. В пероксисомах гликолевая к˝ислота окисляется с участием фермента гликолатоксидазы и превращается в глиоксиловую кислоту:
301

Продукт данной реакции Н2Î2 разлагается под действием каталазы на воду и кислород, а глиоксиловая кислота аминируется от глутаминовой кислоты, превращаясь в аминокислоту глиц˝ин:
Аминокислота глицин не накапливается в пероксисомах, а транспортируется из них в митохондрии, где участвует в си˝нтезе
аминокислоты серина. Эту реакцию катализируют ферменты глициндекарбоксилаза è серинтрансгидрооксиметилаза, содержащая в
активном центре в качестве кофермента тетрагидрофолиев˝ую кис-
лоту. В ходе реакции синтеза серина происходит также высв˝обождение СО2 è NH3, а также образование НАД · Н:
Образовавшийся в митохондриях серин может далее транспо˝р-
тироваться в пероксисомы и под действием аминотрансферазы ïå-
редавать аминогруппу на молекулы пировиноградной кисло˝ты. В
результате этой реакции серин превращается в гидроксипи˝ровиноградную кислоту, а пировиноградная кислота — в аминокис˝лоту аланин:
Гидроксипировиноградная кислота восстанавливается в гл˝ицериновую кислоту с участием дегидрогеназы:
302
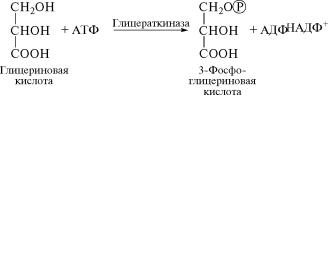
Продукт этой реакции глицериновая кислота может затем в
хлоропластах фосфорилироваться и, превращаясь в 3-фосфогл˝и-
цериновую кислоту, включаться в реакции цикла Кальвина:
Таким образом, при взаимодействии пероксисом, хлороплас-
тов и митохондрий в фотосинтезирующих клетках растений о˝существляется процесс, связанный с поглощением О2 и высвобож-
дением СО2, который называют фотодыханием. Кислород прини-
мает участие в первой реакции, где он связывается вместо С˝Ос
2
молекулами рибулозо-1,5-дифосфата, и при окислении фосфогли˝-
колевой кислоты в пероксисомах. Выделение СО2 происходит в
митохондриях в ходе синтеза аминокислоты серина.
В связи с тем что при фотодыхании осуществляются превраще˝-
ния гликолевой кислоты — продукта разложения первичног˝о акцептора СО2, синтезируемого в реакциях цикла Кальвина, за счет таких превращений уменьшается количество связанной в хо˝де фотосинтеза СО2, в результате чего понижается урожайность расте-
ний. В опытах установлено, что в естественных условиях про˝изра-
стания при повышенных температурах, снижающих концентра˝- цию СО2 в хлоропластах, продуктивность растений вследствие интенсивного фотодыхания может снижаться на 30—40 %.
Исходя из этих данных, учеными-биохимиками сформулирова-
на важнейшая задача для селекционеров и генетиков — соз˝дание новых сортов сельскохозяйственных культур с пониженной˝ ско-
ростью фотодыхания. Одним из главных направлений такой ра˝боты является оптимизация структуры каталитического цент˝ра фер-
мента рибулозодифосфаткарбоксилазы, направленная на ус˝иле-
303
ние карбоксилазной и ослабление оксигеназной активност˝и этого фермента. Для решения указанной проблемы большие надежды˝ возлагаются на применение методов генетической и белков˝ой инженерии.
9.1.4. АССИМИЛЯЦИЯ СО2 Ó Ñ4-РАСТЕНИЙ
После открытия реакций цикла Кальвина различные исследо˝-
ватели изучали фотозависимое включение меченного 14С углерода
ÑÎ2 в первичные продукты фотосинтеза у разных групп расте-
ний. В ходе этих исследований было выяснено, что у зеленых в˝о-
дорослей и в листьях большинства растений умеренной зоны˝ первичным продуктом фотосинтеза, в который происходит ак˝-
тивное включение меченного 14С углерода СО2 при коротких
световых экспозициях, является С3-продукт — 3-фосфоглицери-
новая кислота, в связи с чем такие растения было предложен˝о
называть С3-растениями. Наряду с этим у многих растений тропического происхождения радиоактивный углерод СО2 сначала
включается в С4-продукты (щавелевоуксусная, яблочная, аспара-
гиновая кислоты), поэтому их называют С4-растениями. К ти-
пичным С4-растениям относятся кукуруза, сахарный тростник,
сорго, просо, амарант, лебеда, некоторые виды рода Euphorbia, многие тропические злаки.
Последовательность биохимических реакций, связанная с и˝с-
пользованием СО2 для синтеза дикарбоновых кислот в листьях
Ñ4-растений, впервые была представлена как циклический про-˝
цесс австралийскими учеными М. Д. Хетчем и К. Р. Слэком и п˝о- лучила название цикла Хетча—Слэка.
Ñ4-растения отличаются от С3-растений строением листа. Лис-
òüÿ Ñ4-растений пронизаны сетью сосудисто-волокнистых пучков,
которые окружены плотным слоем клеток обкладочной парен˝химы, а вокруг этих клеток расположен рыхлый слой мезофилла˝. В клетках обкладочной паренхимы находятся крупные хлороп˝ласты, почти не образующие гран; в них локализованы фермент рибу˝ло-
зодифосфаткарбоксилаза и другие ферменты, катализирующ˝ие ре-
акции цикла Кальвина, а в клетках мезофилла листа содержа˝тся типичные для фотосинтезирующих тканей хлоропласты, в кот˝о- рых происходят фотохимические реакции и осуществляется˝ синтез НАДФ · Н и АТФ.
Первичное связывание СО2 и включение его в состав органи- ческого вещества у С4-растений происходит в цитоплазме клеток
мезофилла с участием фермента фосфопируваткарбоксилазы
(4.1.1.31). Первичным акцептором СО2 служит фосфоенолпирови-
ноградная кислота. В ходе реакции образуются щавелевоукс˝усная кислота и неорганический фосфат:
304

(1)
В опытах показано, что непосредственно с первичным акцеп-˝
тором взаимодействует не СО2, а бикарбонат-ионы (НСО3–). Карбоксилирующий фермент фосфопируваткарбоксилаза предст˝авля-
ет собой белок с высокой молекулярной массой (~400 тыс.), вклю˝-
чающий 12 полипептидных субъединиц, с каждой из которых свя˝-
зан катион Mn2+. Молярная активность этого фермента во много
раз выше карбоксилирующей активности фермента цикла Кал˝ь- вина рибулозодифосфаткарбоксилазы, поэтому он способен˝ эф-
фективно катализировать образование щавелевоуксусной к˝ислоты
при низких концентрациях СО2, которые характерны для расте-
ний тропической зоны вследствие ухудшения растворимост˝и СО
2
в физиологической среде при повышенных температурах. Кро˝ме того, фосфопируваткарбоксилаза не взаимодействует с кис˝лоро-
дом и поэтому не может участвовать в фотодыхании, снижающ˝ем
выход фотосинтетических продуктов.
Синтезированная в цитоплазме клеток мезофилла щавелево˝ук-
сусная кислота затем поступает в хлоропласты и с участием˝ восстановленных динуклеотидов НАДФ · Н, являющихся продуктами
световой стадии фотосинтеза, превращается там в яблочную˝ кис-
лоту. Данную реакцию катализирует фермент малатдегидрогеназа:
(2)
Одновременно с превращением в яблочную кислоту щавелевоуксусная кислота может вступать в реакцию переаминирова˝ния с образованием аспарагиновой кислоты:
305

Затем яблочная кислота и аспартат диффундируют из клеток˝ мезофилла в клетки обкладочной паренхимы (рис. 9.5). В хлоро-˝
пластах этих клеток яблочная кислота подвергается окисл˝ительно-
му декарбоксилированию с участием фермента малатдегидрогеназы декарбоксилирующей (1.1.1.40) с образованием пировиноградной
кислоты и СО2:
(3)
Образовавшиеся в этой реакции СО2 и НАДФ · Н далее уча-
ствуют в реакциях цикла Кальвина, которые активно происхо˝дят в хлоропластах клеток обкладочной паренхимы. Чем интенсив˝нее в
них осуществляется декарбоксилирование, тем больше обра˝зуется
ÑÎ2 и НАДФ · Н и, следовательно, тем больше СО2 связывается с
первичным акцептором и восстанавливается до триоз в ходе˝ реак-
ций цикла Кальвина. Продукт декарбоксилирования яблочно˝й кислоты — пировиноградная кислота не используется в ци˝кле
Кальвина, а диффундирует в клетки мезофилла листа, в хлоро˝пла-
стах которых она снова превращается в первичный акцептор˝
ÑÎ2 — фосфоенолпировиноградную кислоту. Эту реакцию катализирует фермент пируватдикиназа (2.7.1.40), который способен за счет гидролиза макроэргической связи АТФ осуществлят˝ь фосфорилирование с участием неорганического фосфата:
(4)
У некоторых С4-растений (амарант, лебеда) декарбоксилирова-
ние яблочной кислоты происходит в митохондриях клеток об˝кла-
дочной паренхимы, и тогда в качестве биоэнергетического п˝ро-
дукта образуются восстановленные динуклеотиды НАД · Н.
306
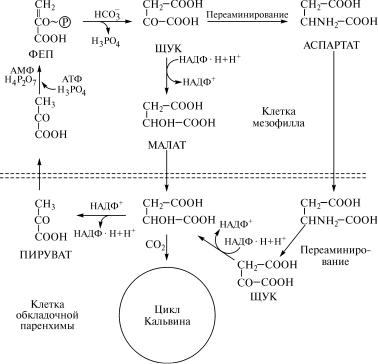
Рис. 9.5. Взаимодействие цикла Хетча—Слэка и цикла Кальвина˝ у С-растений:
4
ФЕП — фосфоенолпировиноградная кислота; ЩУК — шавелев˝оуксусная кислота; МАЛАТ — яблочная кислота; АСПАРТАТ — аспарагиновая кислота; ПИР˝УВАТ — пировиноградная
кислота
Аспарагиновая кислота, синтезируемая в клетках мезофилл˝а
листа, в хлоропластах обкладочной паренхимы вступает в ре˝акции переаминирования, в ходе которых превращается в яблочную˝ кислоту, а яблочная кислота далее подвергается превращениям˝, указанным в реакциях (3) и (4).
Таким образом, у С4-растений наблюдается разделение в про-
странстве биохимических процессов первичного связывани˝я СО
2
и образования продуктов темновой стадии фотосинтеза. Свя˝зывание СО2 происходит в клетках мезофилла листьев в ходе реакций
цикла Хетча—Слэка, и оно сопряжено с потреблением продукт˝ов
световой стадии фотосинтеза НАДФ · Н (при восстановлении ща-
велевоуксусной кислоты в яблочную) и АТФ (в ходе образован˝ия
первичного акцептора СО2 — фосфоенолпировиноградной кисло-
307
ты из пировиноградной). Образование же продуктов темново˝й стадии фотосинтеза осуществляется в клетках обкладочно˝й парен-
химы, в которых функционирует цикл Кальвина, а донорами СО˝
2
являются продукты цикла Хетча—Слэка (яблочная и аспараги˝новая кислоты), подвергающиеся в клетках обкладочной парен˝химы
декарбоксилированию и создающие высокую концентрацию С˝О
2
âэтих клетках.
Âходе декарбоксилирования яблочной кислоты в клетках об˝- кладочной паренхимы синтезируются также восстановленны˝е ди-
нуклеотиды НАДФ · Н (или НАД · Н у некоторых растений). Сл˝е-
довательно, основное назначение цикла Хетча—Слэка — св˝языва-
íèå ÑÎ2 с помощью фермента фосфопируваткарбоксилазы и пе-
ренос его в виде яблочной и аспарагиновой кислот из клето˝к
мезофилла в клетки обкладочной паренхимы, где происходит˝ выс-
вобождение и создание высокой концентрации СО2, необходимой
для более эффективной работы фермента цикла Кальвина риб˝уло-
зодифосфаткарбоксилазы.
Одновременно с транспортом СО2 осуществляется также пере-
нос из клеток мезофилла в клетки обкладочной паренхимы и ˝вос-
становительного потенциала, так как при образовании ябло˝чной кислоты в клетках мезофилла потребляются восстановленн˝ые
динуклеотиды НАДФ · Н, которые снова синтезируются при де-
карбоксилировании яблочной кислоты в клетках обкладочн˝ой паренхимы. Дополнительная энергия в виде молекул АТФ затрач˝и-
вается в клетках мезофилла только на регенерацию первичн˝ого
акцептора СО2 — фосфоенолпировиноградной кислоты, что и определяет в целом дополнительные энергетические затраты˝ на функ-
ционирование реакций цикла Хетча—Слэка. Эти дополнитель˝ные
затраты энергии составляют у С4-растений около 15 % всего коли- чества энергии, потребляемого растениями для образовани˝я фотосинтетических продуктов.
Однако, если учитывать, что С4-растения обычно произрастают
âзасушливой зоне с высокой интенсивностью солнечной рад˝иации, дополнительные затраты на функционирование цикла Хе˝т- ча—Слэка у них не лимитированы солнечной энергией и поэто˝му не снижают эффективности фотосинтеза. Наоборот, у этих ра˝сте-
ний более интенсивно происходит связывание СО2 благодаря действию фермента фосфопируваткарбоксилазы, который не вза˝имодействует с кислородом и, следовательно, не инициирует ре˝акции
фотодыхания, снижающие продуктивность фотосинтеза. И это˝т
фермент имеет очень сильно выраженную субстратную специ˝фич-
ность по отношению к СО2, поэтому способен эффективно связывать СО2 даже при низких его концентрациях, тогда как при таких
условиях карбоксилирующая активность фермента рибулозо˝ди-
308
фосфаткарбоксилазы у С3-растений очень сильно подавляется. Максимальная скорость включения СО2 в органические продукты
ó Ñ4-растений может достигать 40—60 мг на 1 см2 поверхности ли-
ñòà, ÷òî â 1,5 ðàçà âûøå, ÷åì ó Ñ3-растений.
Исходя из указанных выше преимуществ перед С3-растениями, С4-растения имеют более высокий коэффициент использования˝
солнечной энергии и поэтому способны давать более высоки˝й вы-
ход биомассы на единицу возделываемой площади сельскохо˝зяйственных угодий. Наибольшие преимущества перед С3-растения-
ми они имеют в засушливых регионах, так как способны эффек˝-
тивно связывать СО2 даже при почти закрытых устьицах листьев, когда в них сильно понижается концентрация СО2. В более про-
хладных и влажных регионах, где интенсивность солнечной р˝ади-
ации понижена, С3-растения получают определенные преимущества перед С4-растениями, так как им не требуется дополнитель-
ная энергия для фиксации СО2, связанная с функционированием
цикла Хетча—Слэка.
У суккулентных растений (сем. Сrassulaceae), которые также хорошо адаптированы к засушливым условиям, биохимически˝е
процессы связывания СО2 и включения его в реакции цикла Каль-
вина разграничены во времени. Поскольку у этих растений д˝нем
устьица закрыты и нет поступления СО2 из окружающей атмосфе-
ры, поглощение и связывание его происходит ночью под дейс˝твием фермента фосфопируваткарбоксилазы, локализованного ˝в ци-
топлазме клеток листа. Под действием этого фермента СО2 присо-
единяется к фосфоенолпировиноградной кислоте, и таким об˝ра-
зом осуществляется синтез щавелевоуксусной кислоты, кот˝орая
затем восстанавливается в яблочную кислоту. Последняя ко˝нцент-
рируется в вакуолях клеток листа. Первичный акцептор СО2 фосфоенолпировиноградная кислота образуется в результате ˝расщеп-
ления крахмала и осуществления реакций пентозофосфатно˝го
цикла (см. раздел 9.3). Крахмал же накапливается днем, когда и˝н- тенсивно происходят поглощение солнечной энергии и синт˝ез НАДФ · Н и АТФ, необходимых для функционирования цикла Кальвина. Продукты этого цикла активно используются для о˝бра-
зования фотосинтетического крахмала.
Источником СО2 для функционирования цикла Кальвина в
дневное время служит яблочная кислота, которая накаплива˝ется в вакуолях ночью. Она диффундирует в цитоплазму клеток и по˝д-
вергается там декарбоксилированию под действием декарб˝окси-
лирующей малатдегидрогеназы и высвобождает СО2, который поступает в хлоропласты и связывается там под действием фер˝мента рибулозодифосфаткарбоксилазы, а затем подвергается восстановлению до уровня сахаров в реакциях цикла Кальвина. В резул˝ьтате
309
декарбоксилирования яблочной кислоты образуется также ˝пировиноградная кислота, которая под действием фермента пиру˝ват-
дикиназы фосфорилируется и превращается в фосфоенолпир˝ови-
ноградную кислоту. В дальнейшем фосфоенолпировиноградн˝ая кислота через ряд промежуточных реакций (см. раздел 9.3) пре-˝ вращается в триозофосфаты, которые затем участвуют в синт˝езе
глюкозо-6-фосфата, используемого для образования фотосинт˝ети-
ческого крахмала.
9.1.5. ЭФФЕКТИВНОСТЬ ИСПОЛЬЗОВАНИЯ ЭНЕРГИИ ПРИ ФОТОСИНТЕЗЕ
Как было показано ранее, в ходе фотохимических реакций дл˝я образования одной молекулы О2 пигментными комплексами фо-
тосистем I и II поглощается восемь квантов света. Поглотив один
квант красного света, молекулы хлорофилла в реакционных ц˝ентрах фотосистем переходят в первое синглетное возбужденн˝ое со-
стояние, из которого энергия возбуждения может быть испол˝ьзо-
вана для осуществления фотохимических реакций, связанны˝х с инициированием транспорта электронов в мембранах хлоро˝плас-
тов. Если же поглощается квант синего света, обладающего з˝начи-
тельно большей энергией, то молекулы хлорофилла переходя˝т во второе синглетное возбужденное состояние, из которого — в˝ пер-
вое синглетное возбужденное состояние, потеряв часть эне˝ргии в
виде теплоты. Поэтому при поглощении как красного, так и си˝- него света полезное использование энергии происходит то˝лько
из первого синглетного возбужденного состояния и, следовательно, фотосинтетическая активность одного кванта красного˝ и синего света одинакова. Учитывая, что энергия 1 моля красного˝ света (1 эйнштейн) в среднем равна 176 кДж, можно рассчитать общее количество поглощенной энергии для образования 1 мо˝ля О2 : 176 кДж · 8 молей красного света = 1408 кДж.
Из суммарного уравнения фотосинтеза видно, что при образо˝- вании 1 моля О2 происходит поглощение 1 моля СО2, на восстановление которого до уровня фруктозо-6-фосфата в реакциях˝ цикла Кальвина затрачивается 2 моля НАДФ · Н и 3 моля АТФ. Исходя из этого, можно определить энергетические затраты˝ на ассимиляцию хлоропластами 1 моля СО2. Изменение свободной
энергии при синтезе 1 моля АТФ при стандартных условиях со˝-
ставляет 30,6 кДж, а 3 молей АТФ — 91,8 кДж. При восстановле-
íèè 1 ìîëÿ ÍÀÄÔ+ до уровня НАДФ · Н изменение свободной энергии при стандартных условиях равно 219,8 кДж, тогда как
2 молей НАДФ+ — 439,6 кДж. Общее количество энергии, которое
310