

Экспрессия генов Патрушев
.pdf51
которые образуются и в растворах с физиологическими концентрациями солей ( 100 мМ NaCl), содержащих изолированный хроматин. Такие фибриллы в форме соленоида неравномерны по своей длине. С помощью электронного микроскопа в них обнаруживаются гранулы диаметром 30 нм, между которыми находятся более тонкие участки. Ю.С. Ченцов, впервые обнаруживший эти гранулы, назвал их нуклеомерами, позднее они были описаны в литературе под названием "супербидов" (super-beads). Согласно модели А. Клуга, на один шаг (10 нм) соленоида приходится 6 нуклеосом, что приводит к уменьшению в ~6 раз длины фибриллы диаметром в 10 нм, а исходной свободной ДНК – в 40 раз. Таким образом, в соответствии с этой упрощенной картиной цепь нуклеосом на втором уровне упаковки хроматина свернута в симметрично построенный соленоид, содержащий нуклеомеры. Симметрия соленоида нарушается при взаимодействии с ним негистоновых белков.
Энергия, необходимая для образования соленоида, частично черпается из энергии взаимодействия нуклеосом друг с другом. Однако основной вклад в этот процесс вносит гистон Н1, связывание молекул которого с линкерной ДНК за пределами кóровых частиц обеспечивает образование компактных фибрилл диаметром 30 нм. Дифференцирующиеся клетки высших эукариот обладают способностью к синтезу целого семейства гистонов Н1, например Н1А, Н1В, Н1о, которые имеют структурное сходство с гистоном Н5, обнаруженным в дифференцирующихся предшественниках эритроцитов. Полипептидные цепи гистонов семейства Н1, как и молекулы гистонов кóровых частиц, содержат центральный глобулярный домен и два гибких плеча. Глобулярный домен определяет локализацию гистона Н1 в нуклеосомах, где он взаимодействует с ДНК в месте ее входа в кóровую частицу и выхода из нее, стабилизируя два отрицательных супервитка ДНК, обернутой вокруг гистонового октамера. Расщепление хроматина нуклеазой микрококков приводит к освобождению гистона Н1, так как при этом происходит отщепление сегментов ДНК, с которыми он взаимодействует.
Следует подчеркнуть, что точная структура соленоида в настоящее время неизвестна. Данные, недавно полученные методом рассеивания нейтронов, указывают на то, что гистон Н1 локализован внутри хроматиновой фибриллы и на ее поперечном срезе помещается шесть нуклеосом, что хорошо
52
соответствует модели Клуга. Однако при изучении хроматина с помощью сканирующей микроскопии было установлено, что при низкой ионной силе соленоид существует в виде нерегулярной спирали.
Петельно-доменный уровень компактизации хроматина. В
интерфазных ядрах эукариот нити хроматина, в которых ДНК упакована в форме соленоида, содержащего нуклеомеры, организованы в виде топологически независимых петель, длина которых в среднем составляет 50– 100 т.п.о. Такой способ пространственной укладки нитей хроматина рассматривается как следующий уровень конденсации хроматина (и ДНК) у эукариот, а сами петли получили название хромомеров. С помощью электронного микроскопа обнаружено, что нити хроматина в хромомерах имеют дополнительную специфическую укладку в виде розеток, собранных у основания, от которого отходят малые петли длиной ~5 т.п.о. (см. рис. I.2). Образование хромомеров становится возможным благодаря наличию у их оснований определенных последовательностей нуклеотидов, которые специфически взаимодействуют с ядерным матриксом, называемым иначе ядерным скэффолдом (скелетом) – сетчатообразной структурой внутри интерфазных ядер, образованной белками и РНК. Эти участки хромосомной ДНК, взаимодействующие с ядерным матриксом, в литературе известны под сокращенными названиями MAR (Matrix Associated Region) или SAR (Scaffold Associated Region) и часто обозначаются как MAR/SAR-последовательности.
Были предприняты попытки связать локализацию MAR/SARпоследовательностей на ДНК с функциональной активностью хроматина. Исследование петельной структуры хроматина в ядрах дрозофилы показало, что эти последовательности часто располагаются в окрестностях генов, транскрибируемых РНК-полимеразой II, и фланкируют участки хроматина, содержащие одну или несколько транскрипционных единиц. Выяснилось, что активно транскрибируемые гены организованы в петли небольшого размера ( 10 т.п.о.), тогда как "молчащие" гены находятся преимущественно в составе более крупных петель, которые содержат несколько транскрипционных единиц и, в свою очередь, образуют дополнительные петли в виде розеток (см. рис. I.2). Как правило, MAR/SAR-последовательности фланкируют гены, однако в ряде случаев их обнаруживают и внутри генов, но в составе интронов. В настоящее время выделено и охарактеризовано несколько десятков MAR/SAR-
53
последовательностей из генома дрозофилы и других организмов. У дрозофилы они представляют собой АТ-богатые последовательности нуклеотидов длиной в 200–350 п.о., в составе которых часто встречаются участки поли(dA) поли(dT). Обнаружены два типа блоков из АТ-богатых последовательностей: А-блок –
AATAAAT/CAAA и Т-блок – TTA/TTT/ATTT/CAAA, характерные для всех
MAR/SAR-последовательностей не только дрозофилы, но и других организмов. Отличающиеся от них блоки похожих последовательностей нуклеотидов – ATATTT и AATATTTT, первоначально найденные в MAR/SARпоследовательностях мышей, оказались широко представленными и в аналогичных последовательностях многих эукариот.
Можно выделить следующие обобщенные характеристики MAR/SARпоследовательностей, приведенные в недавнем обзоре М.В. Глазкова (1995 г.):
длина MAR/SAR-последовательностей составляет 300–1000 п.о.; все они содержат многочисленные сайты ДНК-белкового взаимодействия и обогащены AT-парами;
MAR/SAR-последовательности обладают способностью обратимо связываться с ядерным матриксом метафазных хромосом и локализованы исключительно в некодирующих последовательностях генома, главным образом в нетранскрибируемых участках, реже в интронах;
расстояние между двумя соседними MAR/SAR-последовательностями составляет 3–112 т.п.о., и они фланкируют один или несколько экспрессирующихся генов, для которых характерна повышенная чувствительность к нуклеазам из-за деконденсированного состояния их хроматина;
некоторые MAR/SAR-последовательности выявлены рядом с энхансероподобными регуляторными элементами. Кроме того, эти последовательности могут быть потенциальными сайтами инициации репликации и содержать в своем составе большое число сайтов для разных факторов транскрипции;
MAR/SAR-последовательности иногда соседствуют с последовательностями, способными образовывать Z- или H-формы ДНК, и в таких случаях они определяются как сайты с повышенной чувствительностью к ДНКазе I.
Все это указывает на то, что MAR/SAR-последовательности являются
54
чрезвычайно важными в функциональном отношении последовательностями геномной ДНК эукариот. Одной из основных функций, которую им приписывают
внастоящее время, может быть пространственное разграничение функциональных доменов хроматина в интерфазных ядрах эукариот, необходимое для эффективной и независимой экспрессии генов, находящихся
вэтих доменах. Подробнее о выполнении MAR/SAR-последовательностями функций пограничных последовательностей (инсуляторов), регулирующих экспрессию эукариотических генов, см. в разделе 3.2.4.
Являясь одними из ключевых цис-действующих генетических регуляторных элементов хромосом эукариот, MAR/SAR-последовательности осуществляют глобальный контроль изменений структуры хроматина и связанных с ними модуляций экспрессии генов на уровне транскрипции. В настоящее время считается, что эти последовательности обеспечивают поддержание функциональной структуры хромосом в интерфазных ядрах и вовлечены в процесс их конденсации при формировании метафазных хромосом во время митоза. Современная модель структуры метафазной хромосомы К.М. Харта и У.К. Лэммли (1998 г.) подчеркивает, что SARпоследовательности, пространственно примыкая друг к другу, образуют ось хроматиды, от которой в разные стороны отходят петли хроматина, формирующие тело метафазной хромосомы. Однако еще остается доказать, являются ли MAR/SAR-последовательности местами специфического прикрепления белковых комплексов, включающих, к примеру, белок конденсин, которые необходимы для конденсации хроматина.
Негистоновые белки хроматина. Большое влияние на структуру хроматина и функционирование эукариотических генов оказывают различные негистоновые белки. В ядрах в наибольшем количестве обнаруживают негистоновые белки, которые относят к так называемой группе белков с высокой подвижностью (high mobility group – HMG). Это название отражает их высокую подвижность при электрофорезе. Суммарное содержание HMG-белков
вядрах клеток в ~10 раз меньше, чем гистонов. Эти белки разделяют на три основных подкласса: 14/17, 1/2 и I/Y.
HMG 14/17 представляют собой небольшие белки с молекулярной массой 10–12 кДа, полипептидные цепи которых несут большой локальный электрический заряд при физиологических значениях рН. N-Концевые части их
55
полипептидных цепей обладают основными свойствами, тогда как С-концевые
– кислыми. Кóровые частицы нуклеосом содержат на своей поверхности две молекулы HMG 14/17, которые соединяют между собой цепи ДНК двух соседних витков. HMG-Белки этой группы могут замещать гистон H1 в активно транскрибируемых генах. In vivo ~10% коровых частиц нуклеосом могут содержать белки HMG 14/17.
Белки группы HMG 1/2 также являются одними из преобладающих ядерных белков с молекулярной массой 25–30 кДа. Эти белки специфически связываются с одноцепочечными участками ДНК, а также участками ДНК, образующими крестообразные структуры в местах, содержащих палиндромные последовательности. Это указывает на их возможное участие в процессах рекомбинации, репарации и(или) репликации. Было показано, что белки HMG 1/2 способствуют сборке нуклеосом in vitro, однако физиологическое значение этого факта остается непонятным, поскольку многие отрицательно заряженные молекулы в экспериментальных условиях также ускоряют образование нуклеосом.
Белки HMG I/Y преимущественно взаимодействуют с АТ-богатыми участками ДНК, что характерно и для гистона Н1. Предполагается, что эти белки конкурируют с гистоном Н1 in vivo за промоторы и области начала репликации ДНК (см. разделы 2.1 и 4.1).
Помимо HMG-белков к негистоновым белкам хроматина относятся многочисленные внутриядерные ферменты и белковые факторы, необходимые для работы генетического аппарата клетки. Многим из этих белков, обеспечивающим транскрипцию, репарацию и репликацию, посвящены целые разделы этой книги. Среди негистоновых белков особое место занимают ДНКтопоизомеразы, механизмы функционирования которых целесообразно рассмотреть отдельно.
1.3.3. Роль ДНК-топоизомераз в обеспечении структуры и функционирования хроматина
Топоизомеразы контролируют в клетках уровень суперскрученности ДНК, который может изменяться в процессе ее репликации, транскрипции, гомологичной рекомбинации, а также во время перестроек хроматина. Все эти
56
ферменты релаксируют суперскрученные молекулы ДНК, снимая их внутреннее напряжение путем внесения одноили двухцепочечных разрывов с последующим их восстановлением (лигированием). По механизму действия различают ДНК-топоизомеразы типа I и II. ДНК-топоизомеразы I, которые являются мономерными белками,
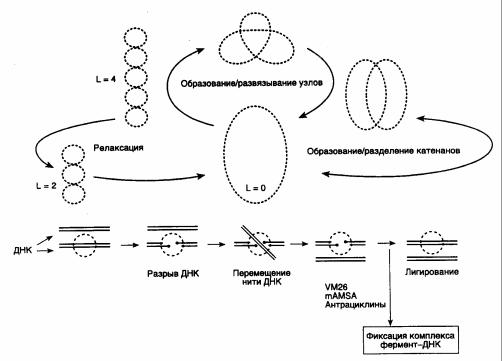
Рис. I.3. Основные этапы каталитического цикла топоизомеразы II
Внизу изображен механизм переноса нити ДНК через двухцепочечный разрыв. Точками отмечены места ковалентного присоединения фермента к 5'-концам ДНК в двухцепочечных разрывах. VM26, mAMSA, антрациклины – ингибиторы топоизомеразы II, фиксирующие ковалентный комплекс фермент-ДНК и предотвращающие лигирование двухцепочечного разрыва; L – число зацеплений ДНК в суперскрученной молекуле.
релаксируют ДНК без затраты энергии путем внесения одноцепочечных разрывов. В отличие от этого, ДНК-топоизомеразы II функционируют в виде димеров, осуществляя ATP-зависимое расщепление обеих цепей ДНК с последующим переносом цепей через разрыв и его лигированием (рис. I.3).
Для внесения одноцепочечного разрыва в ДНК все ДНК-топоизомеразы используют остаток Tyr, который осуществляет нуклеофильную атаку фосфатной группы ДНК с образованием фосфотирозина. В результате
57
ферменты оказываются ковалентно связанными с 5'- или 3'-концами ДНК в одноцепочечном разрыве. Образование такой ковалентной связи исключает необходимость затраты энергии при восстановлении фосфодиэфирной связи в одноцепочечном разрыве на заключительных стадиях реакции. У ДНКтопоизомераз типа I имеется один каталитический остаток Tyr на молекулу мономерного белка, тогда как димеры ДНК-топоизомераз II содержат по одному каталитическому остатку на каждую субъединицу, что обеспечивает создание ступенчатого двухцепочечного разрыва в молекуле релаксируемой ДНК.
Обнаружены, по крайней мере, два подтипа ДНК-топоизомераз I – IA и IB, которые, будучи неродственными ферментами как по первичной, так и по пространственной структурам, выполняют аналогичные функции с помощью различных механизмов. До недавнего времени ДНК-топоизомеразы IA считали исключительно прокариотическими ферментами, однако они были найдены и в клетках эукариот, включая клетки человека, и названы ДНКтопоизомеразами III.
ДНК-топоизомераза IA релаксирует ДНК, содержащие только отрицательные супервитки, работает в присутствии ионов Mg2+ и ковалентно соединяется с 5'-концами ДНК в образующихся врéменных одноцепочечных разрывах. Это сближает ДНК-топоизомеразы IA и II между собой, что было подтверждено также структурными исследованиями. В отличие от этого, ДНКтопоизомеразы IB способны релаксировать ДНК как с положительными, так и отрицательными супервитками, не требуют для своего функционирования ионов металлов и взаимодействует ковалентно с 3'-концами ДНК. ДНКтопоизомеразы IB найдены исключительно в клетках эукариот (за единственным исключением вируса вакцины).
В клетках человека ДНК-топоизомераза IB/III специфически ингибируется камптотецином (camptothecin), который в настоящее время рассматривается в качестве перспективного противоопухолевого препарата. Это соединение взаимодействует преимущественно с ковалентным комплексом топоизомераза I–ДНК, что подавляет реакцию восстановления фосфодиэфирной связи и освобождение фермента из комплекса. В результате происходит быстрое накопление двухцепочечных разрывов ДНК и вступление клеток в апоптоз.
ДНК-топоизомераза II является жизненно важным ферментом любого
58
эукариотического организма. Кроме релаксации суперскрученных молекул ДНК она может осуществлять образование или развязывание узлов, а также образование или разделение катенанов (кольцевых замкнутых ДНК, сцепленных друг с другом). Реакции развязывания узлов и разделения катенанов являются прерогативой именно ДНК-топоизомеразы II и не выполняются ДНК-топоизомеразами I.
У дрожжей ДНК-топоизомераза II требуется для разделения катенанов сестринских хроматид хромосом в анафазе митоза и абсолютно необходима для сегрегации хромосом в мейозе, а также конденсации хроматина в процессе формирования метафазных хромосом. Выяснена важная роль ДНКтопоизомеразы II и в формировании высших уровней структуры хроматина, а именно участие фермента в образовании петель хроматина во время конденсации хромосом.
ДНК-топоизомераза II локализована в ядре и в больших количествах ассоциируется с ДНК как в интерфазных, так и метафазных ядрах. С помощью специфических антител показано, что молекулы фермента располагаются преимущественно вдоль центральной продольной оси обоих плеч хромосом у многих организмов. Такое аксиальное распределение ДНК-топоизомеразы II в хромосомах наблюдали даже после удаления из них большей части гистонов в результате многократных солевых экстракций. Специфическая локализация этого фермента в хромосомах очень показательна в свете обсуждавшихся выше петельно-доменных особенностей организации хроматина в ядрах. Создается впечатление, что ДНК-топоизомераза II находится в виде гомодимера в основании петель, взаимодействуя с MAR/SARпоследовательностями ДНК хроматина. Хотя топоизомераза II не обнаруживает строгой специфичности в отношении расщепляемых последовательностей нуклеотидов, на выбор сайтов большое влияние оказывают структурные компоненты хроматина. Показано, что in vivo существуют два класса сайтов, по которым происходит расщепление ДНК этим ферментом: одни из них локализованы в активно транскрибируемых участках хроматина, гиперчувствительных к действию нуклеаз, а другие – непосредственно в MAR/SAR-последовательностях. Ассоциация ДНК-топоизомеразы II с активно транскрибируемыми участками хроматина указывает на ее возможную важную роль в регуляции экспрессии генов, что и было продемонстрировано в недавних
59
экспериментах. Таким образом, ДНК-топоизомераза II является одним из ключевых ферментов, необходимых для разрешения сложных топологических проблем, возникающих при изменении структуры хроматина в процессах репликации ДНК, транскрипции генов и сегрегации хромосом в митозе и мейозе. Определенные изоформы (α или β) этого фермента, по-видимому, играют важную роль в поддержании динамической структуры хроматина интерфазных и митотических хромосом.
Имеющиеся в настоящее время данные свидетельствуют о высокоупорядоченной организации генома у любого, особенно эукариотического, организма. Первым уровнем такой упорядоченности является консервативное линейное распределение генов и других последовательностей нуклеотидов вдоль молекул ДНК хромосом, которое служит важным таксономическим признаком. Другим не менее жизненно важным свойством генома эукариот и, по-видимому, таксономическим признаком является его упорядоченное распределение в объеме интерфазных ядер. Высокоспецифическое пространственное распределение хроматина эукариот в интерфазных ядрах можно рассматривать в качестве второго уровня его упорядоченности. Находясь в деконденсированном состоянии после завершения митоза, интерфазные хромосомы не перемешиваются внутри интерфазных ядер, но занимают вполне определенные микрокомпартменты. Определенные участки хромосом, маркированные специфическими (в частности MAR/SAR) последовательностями, служат для прикрепления ДНК хромосом к компонентам ядерного матрикса и ядерных мембран. Такие контакты необходимы для эффективной реализации генетической информации в процессе экспрессии генов, эффективной конденсации хроматина и разделения хромосом в митозе и мейозе. Кроме того, пространственно организованное распределение генетического материала в интерфазных ядрах обеспечивает дифференциальную защиту от мутаций отдельных генетических локусов и, по-видимому, может контролировать темп и направление эволюционных изменений как отдельных локусов, так и организмов в целом (подробнее см. раздел 5.3). К сожалению, исследование пространственной организации генома в интерфазных ядрах (архитектоники ядра) сопряжено с большими методическими трудностями и сегодня еще только начинается.
60
ГЛАВА 2. РЕАЛИЗАЦИЯ ГЕНЕТИЧЕСКОЙ ИНФОРМАЦИИ ПРИ ЭКСПРЕССИИ ГЕНОВ
Конечным результатом экспрессии генов, кодирующих белки или нуклеиновые кислоты, должно быть образование этих полноценных в функциональном отношении макромолекул, сопровождаемое формированием определенного фенотипа организма. В соответствии с основным постулатом молекулярной биологии генетическая информация в процессе ее реализации передается однонаправленно от нуклеиновых кислот к белкам. При этом реализуется следующая обобщенная схема: ДНК ↔ РНК → белок, которая подчеркивает, что в ряде специальных случаев возможна передача генетической информации от РНК к ДНК с использованием механизма обратной транскрипции. До сих пор не обнаружена передача генетической информации от белков к нуклеиновым кислотам. На первом этапе экспрессии генов происходит переписывание генетической информации, заключенной в генах, на матричные (информационные) РНК (мРНК – messenger RNA, mRNA), которые являются местом промежуточного хранения этой информации при ее реализации. В некоторых случаях уже сами РНК являются конечным результатом экспрессии генов, и после ряда ферментативных модификаций они непосредственно используются в клеточных процессах. Это относится, прежде всего, к рибосомным и транспортным РНК (рРНК и тРНК), которые вместе составляют основную часть суммарной РНК клетки. К таким РНК принадлежат и малые ядерные РНК (мяРНК), участвующие в процессинге предшественников мРНК эукариот, РНК, входящие в состав ферментов, и природные антисмысловые РНК.
Синтез РНК происходит в результате сложной последовательности биохимических реакций, называемой транскрипцией. Появление русифицированного термина "мРНК" связано с тем, что на втором этапе реализации генетической информации, называемом трансляцией, последовательность нуклеотидов мРНК согласно генетическому коду однозначно определяет последовательность аминокислотных остатков синтезируемых белков, т.е. является матрицей, в соответствии с последовательностями нуклеотидов которой происходит соединение