

Экспрессия генов Патрушев
.pdf151
полужизни мРНК в цитоплазме. Без тщательного контроля над этим процессом с помощью селективного деаденилирования невозможно регулировать внутриклеточную деградацию мРНК, а вместе с тем и уровень экспрессии соответствующих генов с участием данного механизма.
Полиаденилирование является универсальным феноменом, играющим важную роль в процессинге и функционировании мРНК как прокариотических, так и эукариотических организмов. Однако сравнение механизмов полиаденилирования у этих групп организмов выявляет существенные различия, суммированные в табл. I.9.
Н. Саркаром (1997 г.) было высказано предположение о возникновении механизма полиаденилирования РНК. Он полагает, что поскольку у бактериальных и эукариотических ферментов, выполняющих аналогичные функции, не обнаружено гомологии в аминокислотных последовательностях, оба фермента возникли недавно из уже значительно дивергировавших функционально родственных предшественников. Такими предшественниками могли быть прокариотические и эукариотические тРНКнуклеотидилтрансферазы, осуществляющие посттранскрипционный синтез последовательности CCA на 3’-концах тРНК, что по своему механизму близко к полиаденилированию. Подтверждением этого является обнаруженная значительная гомология между тРНК-нуклеотидилтрансферазой и основной поли(А)-полимеразой E. coli, а также между тРНК-нуклеотидилтрансферазой бактерии Sulfolobus shibatae, обитающей в горячих серных источниках, и поли(А)-полимеразами эукариот. Следы такой эволюционной связи обнаруживаются и в современных митохондриях, где в результате полиаденилирования мРНК могут создаваться терминирующие кодоны. По мнению Саркара, различия между системами полиаденилирования прокариот и эукариот, представленные в табл. I.9, можно рассматривать в качестве продукта эволюционной дивергенции сходных биосинтетических функций в процессе независимого возникновения нового регуляторного механизма, обеспечивающего физиологические нужды уже глубоко различающихся групп организмов.

|
|
152 |
|
|
Таблица I.9 |
Сравнение полиаденилирования мРНК у эукариот и прокариот |
||
|
|
|
Функции |
Млекопитающие |
E. coli |
|
|
|
Длина поли(А)- |
80–200 |
14–60 |
последовательностей, нт |
|
|
Уровень |
100 |
2–50 |
полиаденилирования |
|
|
отдельных мРНК, % |
|
|
Локализация сайтов |
Ниже консенсусной |
Любые доступные 3’-OH- |
полиаденилирования |
последовательности |
концы мРНК |
|
AAUAAA |
|
Функции: |
|
|
стабильность мРНК |
Стабилизация мРНК без |
Дестабилизация мРНК, |
|
участия 3’-концевых |
обладающих 3’- |
|
структур типа "стебель– |
концевыми структурами |
|
петля" |
типа "стебель–петля", |
|
|
возможная стабилизация |
|
|
мРНК без таких структур |
трансляция |
Специфический контакт |
Связывание |
|
с белком PABP, |
рибосомного белка S1, |
|
необходимый для |
возможно, необходимое |
|
взаимодействия мРНК с |
для взаимодействия |
|
40S субчастицами |
мРНК с |
|
рибосом |
30S субчастицами |
|
|
рибосом |
репликация плазмид |
Не участвует |
Деградация |
|
|
антисмысловой РНК, |
ингибирующей репликацию плазмид типа ColE1
153
Сплайсинг. Мозаичная интрон-экзонная структура генов эукариот предполагает функционирование механизма, который бы распознавал интроны в предшественниках РНК и с высокой точностью удалял их. Действительно, в 1977 г. такой процесс был обнаружен, он получил название сплайсинга (от англ. splice – соединять концами) и с тех пор интенсивно исследуется.
Удаление последовательностей интронов с помощью сплайсинга происходит в ядрах эукариот сразу после завершения синтеза пре-РНК. В сплайсинге, как правило, участвуют особые рибонуклеопротеиновые (РНП)- частицы – малые ядерные РНП (мяРНП), в состав которых входят мяРНК U1– U6 и многочисленные белки. Эти РНП-частицы на стыках интронов и экзонов образуют функциональный комплекс, получивший название сплайсомы. Однако не все интроны для своего удаления требуют функционирования такого сложного аппарата. В частности, интроны предшественников тРНК у эукариот удаляются с участием более простого набора ферментов, а для вырезания некоторых интронов не требуется никаких дополнительных компонентов, кроме самих предшественников РНК. Последний процесс получил название аутосплайсинга (self-splicing). Поскольку механизмы химических реакций, происходящих в ходе аутосплайсинга, реализуются и при функционировании сплайсомы, они будут рассмотрены более подробно в следующем разделе.
В заключение этого краткого введения определим с помощью приведенной ниже схемы термины, которые будут использоваться при обсуждении механизмов сплайсинга.
5’-Концевой сайт |
Точка |
3’-Концевой сайт |
сплайсинга |
разветвления |
сплайсинга |
↓ |
↓ |
↓ |
5’– экзон 1 GUAUGU__... |
__UACUAAC__... |
__(Py)nAG экзон 2 –3’ |
←----------------------- |
Интрон--------------------- |
→ |
На схеме изображен интрон, соединяющий два соседних экзона в предшественнике мРНК дрожжей. В общих чертах та же структура характерна и для пре-мРНК высших организмов. Места соединения интронов и экзонов, в которых происходит разрыв фосфодиэфирных связей пре-мРНК во время сплайсинга, в зависимости от их положения в интроне называют 5’- или 3’- концевыми сайтами сплайсинга. Полипиримидиновая последовательность
154
(Py)n перед 3'-концевым сайтом сплайсинга существенна для правильного вырезания интронов. Остаток аденозина в консервативной последовательности нуклеотидов интрона, расположенный ближе к его 3’-концу, получил название точки разветвления (branch point). Именно с этим аденозином ковалентно соединяется 5’-конец интрона, освобождающийся на первом этапе сплайсинга с образованием структуры типа "лассо" (lariat) (см. ниже). Первичная структура указанных сайтов мало консервативна в генах, кодирующих ядерные пре-мРНК, и может значительно варьировать даже у интронов одного и того же организма. В зависимости от механизма вырезания интронов и особенностей их пространственной структуры различают интроны групп I, II и III, интроны ядерных РНК, а также твинтроны – интроны, расположенные внутри интронов.
Аутосплайсинг интронов групп I, II и III. Открытие интронов,
удаляющихся из пре-РНК в результате аутокаталитического процесса – аутосплайсинга, имело далеко идущие последствия для развития фундаментальных и прикладных исследований в молекулярной генетике и породило целое направление с использованием рибозимов (подробнее см. раздел 9.2). Такие интроны впервые были обнаружены в органеллах высших организмов, но вскоре было продемонстрировано их широкое распространение в природе. Исследование молекулярных механизмов аутосплайсинга показало, что хотя все вышеупомянутые интроны в сплайсинге используют одни и те же реакции трансэтерификации:
О− О−
R–OH + R’–O–P–OR’’ → R–O–P–OR’’ + R’–OH,
║║
ОО
детали механизмов для интронов групп I, II и III различны.
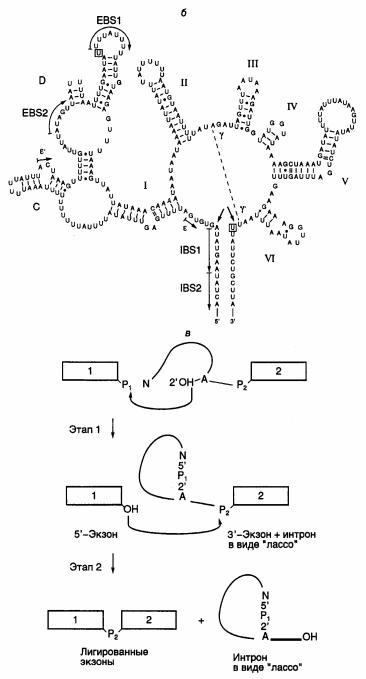
На рис. I.15,а,б изображены структурные особенности интронов I–III групп. Интроны группы I образуют наиболее сложные вторичные и третичные структуры. Они обнаружены в предшественниках РНК простейшего Tetrahymena thermophila и удаляются из них с использованием приведенного в табл. I.10 механизма.

155
Таблица I.10
Механизм прямой и обратной реакций аутосплайсинга интронов группы I
Аутосплайсинг |
Обратное лигирование |
|
|
GTPOH |
GTPOH |
+ |
+ |
[Экзон 1]upA−Интрон−Gpa[Экзон 2] |
[Экзон 1]upA−Интрон−Gpa[Экзон 2] |
↓ |
↑ |
Расщепление |
Интеграция |
5’-концевого сайта |
интрона в |
сплайсинга |
прежнее место |
↓ |
↑ |
[Экзон 1]u−OH |
[Экзон 1]u−OH |
GpA−Интрон−Gpa[Экзон 2] |
GpA−Интрон−Gpa[Экзон 2] |
↓ |
↑ |
Лигирование |
Лигирование |
экзонов |
экзона 2 и интрона |
↓ |
↑ |
[Экзон 1]upa[Экзон 2] |
[Экзон 1]upa[Экзон 2] |
+ |
+ |
GpA−Интрон−G |
GpA−Интрон−G |
|
|

156
157
Рис. I.15. Структура интронов I-III групп и механизм аутосплайсинга
а – вторичные структуры типичных интронов типа I (Tetrahymena thermophila), II и III групп (Euglena gracilis). Стрелками обозначены 3′- и 5′-
сайты сплайсинга, а также места интеграции твинтронов, пунктирными линиями – сайты интронов, сближенные в пространстве, EBS и IBS – соответственно сайты связывания экзонов и интронов; б – предполагаемая вторичная структура интрона группы II; в – двухэтапный
158
механизм аутосплайсинга. Римскими цифрами обозначены предполагаемые двухцепочечные участки РНК, 1, 2 – экзоны
Строчными буквами в таблице изображены нуклеотиды, принадлежащие экзонам, а прописными – интронам. Прямая и обратная реакции не требуют белковых катализаторов. Однако в ряде случаев интроны I и II групп могут кодировать полипептид, названный матуразой (maturase, от англ. mature – созревать), который необходим для эффективного аутосплайсинга и транспозиции интронов in vivo.
Аутосплайсинг интронов группы I начинается с нуклеофильной атаки 2’- OH-группы GTP по фосфодиэфирной связи в 5’-концевом сайте сплайсинга (см. табл. I.10) (присутствие GTP в качестве кофактора необходимо для протекания данной реакции). При этом происходит освобождение 5’-концевого экзона 1. Образовавшиеся половинки молекулы удерживаются рядом друг с другом водородными связями. Освободившаяся 3’-OH-группа экзона 1 далее атакует фосфодиэфирную связь в 3’-концевом сайте сплайсинга, что сопровождается расщеплением этой связи и полным освобождением интрона. Одновременно происходит лигирование двух экзонов. Все этапы этой реакции обратимы. В обратной реакции после связывания соответствующего участка лигированных экзонов "активным центром" вырезанного интрона последний атакует своей концевой 3’-OH-группой фосфодиэфирную связь в месте стыковки экзонов, что сопровождается объединением интрона и экзона 2. На втором этапе 3’-OH- группа экзона 1 атакует фосфатную группу по первой фосфодиэфирной связи интрона, освобождая остаток G, с последующим объединением экзона 1 с остальной частью молекулы.
Аутосплайсинг интронов II и III групп протекает по другому механизму (см. рис. I.15,в). Реакция также осуществляется в два этапа. На первом этапе 2’-OH-группа аденозина, находящегося в точке разветвления интрона, вырезаемого из предшественника РНК, атакует фосфодиэфирную связь 5’- концевого сайта сплайсинга, что сопровождается освобождением экзона 1 и образованием промежуточной структуры в виде лассо. В таком промежуточном соединении 5’-концевой нуклеотид интрона соединен 2’–5’-фосфодиэфирной связью с этим остатком аденозина, образуя петлю на конце интрона. На втором этапе 3’-концевая OH-группа экзона 1 атакует 3’-концевой сайт сплайсинга. В
159
результате происходит объединение экзонов и освобождается интрон с петлей на 5’-конце. В заключение следует отметить одно замечательное свойство этой группы реакций. Поскольку число фосфодиэфирных связей на протяжении всех преобразований остается неизменным, они не требуют затраты дополнительной энергии и протекают самопроизвольно.
Сплайсинг у ядерных пре-мРНК. Исследование механизмов сплайсинга у ядерных пре-мРНК показало, что они существенно не отличаются от только что рассмотренных механизмов, используемых для удаления интронов II и III групп, однако ряд различий имеется.
Структура интронов, удаляемых из предшественников РНК аутосплайсингом, высококонсервативна. У интронов ядерных РНК консервативны лишь короткие последовательности в окрестностях сайтов сплайсинга и точки разветвления. Вторая отличительная особенность сплайсинга у ядерных пре-мРНК – потребность в ATP и мяРНП, формирующих сплайсому. Тем не менее, одинаковые черты этих двух систем сплайсинга более существенны. Малые ядерные РНК, входящие в состав мяРНП и сплайсомы, участвуют в распознавании сайтов сплайсинга и точки разветвления, а также в правильном ориентировании экзонов по отношению друг к другу. Их OH-группы осуществляют атаку фосфодиэфирной связи 5’- концевого сайта сплайсинга на первом этапе реакции. Так же как и в случае интронов II и III групп, для вырезаемого интрона характерна структура типа лассо.
Распознавание канонических сайтов при сплайсинге ядерных пре-мРНК.
Для полного прохождения всех реакций сплайсинга его аппарат (сплайсома) должен распознавать на пре-мРНК три критические последовательности нуклеотидов: 3’- и 5’-концевые сайты сплайсинга и точку разветвления. В соответствии с наиболее распространенной моделью поиска экзонов в премРНК с большими интронами, аппарат сплайсинга, прежде всего, отыскивает два близкорасположенных сайта сплайсинга в нужной ориентации. Распознавание конкретной пары сайтов сплайсинга в пре-мРНК дрожжей начинается с взаимодействия по принципу комплементарности между последовательностями этих сайтов и 5’-концевыми участками мяРНК U1, входящих в состав U1-мяРНП-частиц в случае 5’-концевых сайтов сплайсинга, а также белка U2AF65, находящегося в комплексе с U2AF35, с
160
последовательностью 3’-концевого сайта сплайсинга. Эффективность функционирования сайтов сплайсинга зависит от степени соответствия их первичной структуры консенсусным последовательностям.
Сближение двух сайтов сплайсинга, сопровождаемое выпетливанием последовательности интрона, осуществляется с участием SR-белков, обогащенных Ser и Arg, которые взаимодействуют с 70К-белком (70 кДа), входящим в состав U1-мяРНП-частицы, а также U2AF35 на другом конце интрона. Такие комплексы, образованные с участием SR-белков, получили название "коммитированных комплексов".
В то время как 5’-концевые сайты сплайсинга довольно консервативны, 3’-концевые сайты обнаруживают большую вариабельность. U1-мяРНП, находящиеся в комплексе с 5’-концевым сайтом сплайсинга, стимулируют присоединение белков U2AF к 3’-концевому сайту и способствуют правильному распознаванию соответствующего экзона в процессе, получившем название "определение экзона". Эффективное распознавание сайтов сплайсинга соответствующими белками является критическим этапом в правильном и упорядоченном удалении экзонов из пре-мРНК в процессе конститутивного сплайсинга.
Механизмы определения экзонов. Гены позвоночных, как правило,
содержат большое число мелких экзонов (средняя длина 137 п.о.), разделенных протяженными последовательностями интронов, длина которых может превышать 100 т.п.о. При этом ферментные системы, осуществляющие сплайсинг, должны распознавать небольшие экзоны, затерянные среди гигантских некодирующих последовательностей, и осуществлять их правильное объединение друг с другом. Как уже упоминалось выше, канонические (консенсусные) последовательности, необходимые для распознавания экзонов, располагаются на концах интронов. Однако в отличие от соответствующих последовательностей интронов дрожжей регуляторные последовательности в окрестностях сайтов сплайсинга животных менее консервативны, и это, на первый взгляд, значительно усложняет задачу распознавания экзонов системами сплайсинга. В действительности распознавание индивидуальных сайтов сплайсинга у животных, по-видимому, не связано с независимым распознаванием канонических последовательностей в окрестностях каждого экзона. В соответствии с наиболее популярной в настоящее время моделью