

книги из ГПНТБ / Фрумкин, М. Л. Технологические основы радиационной обработки пищевых продуктов
.pdfспособность вызывать порчу, чем у R. nigricans. Доза 300 крад настолько подавляет жизнедеятельность таких специфичных радиолабильиых представителей порчи яблок, как Monilia fructigena и Penicillium expansum, что они не поражают плоды и че рез 5 сут. Важно, однако, подчеркнуть, что для этих грибов су ществуют своп сублетальные дозы (50—100 крад), после облу чения которыми у них так же, как у радиоустойчивых грибов после облучения дозами 200—300 крад, восстанавливается жиз недеятельность и способность вызывать порчу.
Теперь остановимся на вопросе, который возникает у всех изучающих влпяние ионизирующих излучений на микробиоло гическую порчу плодов. Является ли облученное сырье лучшим субстратом для развития микроорганизмов, если учесть, что под действием радиации разрушаются некоторые сложные химиче ские соединения и образуются простые, легко усвояемые ве щества?
Изучить полпостыо вопрос о действии радиации иа субстрат довольно сложно в связи с тем, что свежие плоды и овощи яв ляются живыми биологическими объектами и изменения в них могут быть самыми разнообразными. Проводить исследования на модельных системах нет смысла, так как даже при достаточном соблюдении соотношения отдельных химических компонентов (сахара, органические кислоты, пектиновые вещества и т. д.) не возможно воссоздать механизм гидролитических процессов, про текающих в плодах при радиационном воздействии.
Учитывая изложенное, мы проследили за развитием порчи об лученных и необлучеппых плодов, зараженных облученными и необлученнымп культурами плесневых грибов. Для этого конт рольные и облученные дозами 100 и 300 крад плоды заражали необлученными и облученными такими же дозами грибами, на нося суспензии спор определенной концентрации или одинако вые кусочки мпцелия на специально поврежденную поверх
ность плода. Плоды заражали сразу |
и через 5—6 дней после |
облучения, когда жизнедеятельность |
гриба восстанавливается, |
и хранили в камерах с влажностью воздуха 90—95%. |
|
На рис. 8 показана динамика распространения порчи па необлученных и облученных яблоках, зараженных необлученным и
облученным грибом R. nigricans. Плоды |
облучали дозой |
|
300 крад. Как видно из рисунка, скорость распространения необ- |
||
лученного гриба на контрольных и облученных плодах практи |
||
чески |
одинакова. На иеоблучениых плодах |
гриб, облученный |
32 дозой |
300 крад, развивался значительно медленнее, чем необлу- |
|
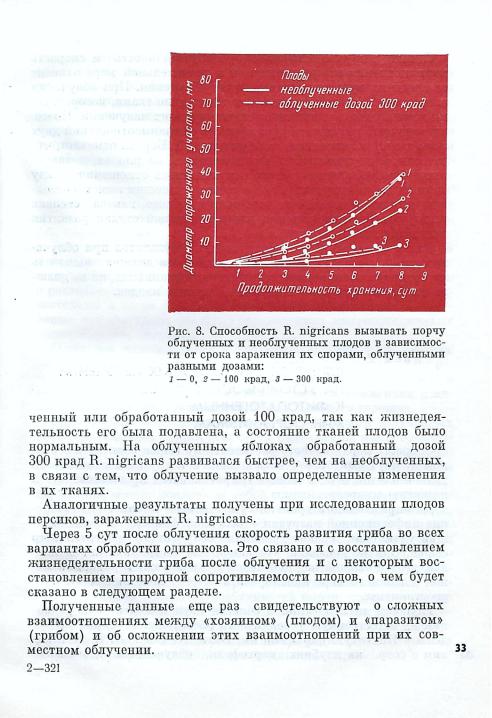
Однако, как уже отмечалось, радиоустойчивость и скорость восстановления микроорганизмов в значительной мере зависят от среды их культивирования после облучения. При облучении плодов такой средой являются растительные ткани, которые са ми изменяются под влиянием ионизирующих излучений. Поэто му конечный результат будет зависеть от взаимоотношений двух живых систем: «хозяина» и «паразита». Л. Бераха отмечает, что латентный период грибов, развивающихся на плодах, в значи тельной мере зависит от того, как сложились отношения между «хозяином» и «паразитом» и в каком физиологическом состоя нии находится каждый из них, точнее от того, какова степень радиационного повреждения плода и в какой стадии развития находится плесневый гриб.
Для получения удовлетворительных результатов при облуче нии плодов, ягод и овощей выбранная доза должна вызывать глубокие изменения в патогенных микроорганизмах, но не долж на оказывать резкого воздействия па ткани плодов.
Г л а в а 2
ВЛИЯНИЕ ИОНИЗИРУЮЩИХ ИЗЛУЧЕНИЙ НА УСТОЙЧИВОСТЬ ПЛОДОВ К ФИТОПАТОГЕННЫМ МИКРООРГАНИЗМАМ
Скорость порчи сочного растительного сырья зависит в основ ном от двух факторов — инфекционной нагрузки и способности тканей плодов, ягод и овощей сопротивляться проникновению и развитию микроорганизмов.
До определенного времени существовало мнение, что сниже ние инфекционной нагрузки — основной фактор, обеспечиваю щий удлинение сроков хранения облученного растительного сырья. Однако доказано, что изменение состояния тканей пло дов, ягод и овощей под действием ионизирующей радиации мо жет в какой-то мере нивелировать эффект действия ее на мик роорганизмы.
Глубокие исследования в области изучения действия радиа ции на естественную устойчивость растительного сырья прове дены в Институте биохимии им. Баха АН СССР Л. В. Метлицким с сотр. на клубнях картофеля, облученных дозами 5—
10 крад для предотвращения прорастания [35, 36]. Авторы при водят обширный материал, свидетельствующий о снижении устойчивости клубней после облучения и вскрывают некоторые причины этого явления. При тщательном изучении всех факто ров, обусловливающих длительность хранения клубней картофе ля в пострадиационный период, они доказали, что снижение устойчивости тканей картофеля под действием радиационной обработки не может быть препятствием для практического ис пользования ее. При соблюдении соответствующих условий (ка чество сырья, продолжительность хранения до облучения, режи мы хранения) облучение картофеля и связанное с ним снижение потерь в результате уменьшения прорастания может дать определеппый эффект.
Все сказанное относится к растительному сырью, в частности к клубням, которые в силу физиологических особенностей мож но и необходимо хранить длительное время. Иначе обстоит дело с растительными объектами, сроки хранения которых непродол жительны и часто определяются днями или неделями. Естест венно, что для таких объектов зараженность микроорганизмами имеет большое значение.
О том, что ионизирующие излучения повышают поражаемость плодов, ягод и овощей микроорганизмами, свидетельствуют ра боты многих исследователей. Отмечено увеличение атакуемости микроорганизмами облученных яблок, томатов, земляники и ма лины, черешни, цитрусовых, винограда, зеленого горошка.
Одни ученые объясняют большую поражаемость облученных объектов стимулирующим действием радиации на микроорганиз мы, другие — лучшей усвояемостью облученного субстрата. Но во всех исследоваипях лишь констатируется факт повышения поражаемости растительного сырья после облучения и отмечает ся, что оно может повлечь за собой изменение сроков хранения
ихарактера порчи. Причины же этого явления не вскрываются.
Влитературе практически отсутствуют и данные о взаимо действии двух факторов — снижения инфекционной нагрузки и изменения природной устойчивости тканей плодов, ягод и ово щей, обработанных ионизирующими излучениями для удлине ния сроков хранения. Некоторые исследователи касаются дей
ствия радиации только на возбудителей порчи, другие — только на растительные объекты. Взаимодействие же этих факторов — одно из главных в комплексе вопросов, решение которых позво ляет правильно оценить перспективность радиационного способа обработки скоропортящегося растительного сырья.
В лаборатории радиобиологии ВНИИКОПа проводились ис следования, в процессе которых необходимо было решить сле дующие задачи:
1.Выявить изменения сопротивляемости плодов инфекции под воздействием различных доз радиации и иитепсивиости из лучений.
2.Определить влияние степени зрелости сырья, вида патоге на, временп внесения инфекции на природную устойчивость плодов, облученных различными дозами ионизирующих излу чений.
3.Изучить возможные причины изменения устойчивости пло дов к микроорганизмам после облучения дозами, рекомендуемы ми для обработки сочного растительного сырья.
Ф А К Т О Р Ы , В Л И Я Ю Щ И Е Н А С О П Р О Т И В Л Я Е М О С Т Ь П Л О Д О В Р А З В И Т И Ю М И К Р О О Р Г А Н И З М О В
Величина л мощность дозы. Из даппых табл. 5 видно, что с возрастанием дозы облучения увеличивается заражаемость пло дов Rhizopus nigricans Ehr. ex Fr. При этом продолжительность инкубационного периода развития возбудителя заболевания со кращается, а скорость его распространения увеличивается.
Отмеченные факты свидетельствуют об ослаблении устойчи вости облученных плодов к микроорганизмам с увеличением дозы облучения.
Заметно ослабляют сопротивляемость незрелых плодов к мик роорганизмам дозы порядка 100 крад. Незначительное действие на ткани оказывают дозы до 50 крад. У некоторых плодов (пер сики) облучение дозой 50 крад вообще не приводит к ослабле нию устойчивости тканей.
Анализ полученных данных показывает, что пострадиацион ное снижение устойчивости плодов является проявлением обще биологической закономерности воздействия радиации на естест венную сопротивляемость растительных объектов микроорганиз мам. При этом у плодов в отличие от вегетирующих растений и запасающих органов растений ослабление устойчивости наблюдается при более высоких дозах облучения.
Так, если у вегетирующих растений снижение сопротивляе мости инфекции отмечается при дозах от 1,5 до 30 крад, а у за пасающих органов — клубней и корнеплодов — при дозах выше
36 7—10 крад, то у плодов — при дозах выше 50—100 крад.
|
|
|
|
Т А Б Л И Ц А 5 |
|
|
|
Количество зара |
Продолжительность |
|
|
Доза |
облучения, |
зившихся плодов |
Скорость |
||
инкубационного |
|||||
крад |
|
через 2 дня после |
периода развития |
распространения |
|
|
|
шюкулнровання |
гриба, ч |
гриба, мы/ч |
|
|
|
гриба, % |
|
|
|
|
|
А б р и к о с ы с о р т а К р а с н о щ е к и й |
|
||
0 |
( к о н т р о л ь ) |
50,0 |
45 |
0,31 |
|
|
50 |
62,5 |
37 |
0,36 |
|
|
300 |
87,5 |
20 |
0,43 |
|
|
|
П е р с п к п с о р т а Э л ь б е р т а |
|
||
0 |
(контроль) |
54,3 |
60 |
0,30 |
|
|
50 |
38,0 |
60 |
0,26 |
|
|
300 |
85,0 |
42 |
0,52 |
|
|
|
Я б л о к и с о р т а А н т о н о в к а о б ы к н о в е н на я |
|||
0 |
(контроль) |
20,8 |
200 |
0,09 |
|
|
50 |
37,0 |
91 |
0,18 |
|
|
300 |
66,5 |
66 |
0,25 |
|
Степень пострадиационного ослабления устойчивости к пара зитам зависит также от мощности дозы облучения. Например, в опытах с яблоками, облученными дозой 300 крад, уменьшение мощности доз с 650 до 250 рад/с способствовало некоторому снижению пострадиационных нарушений, связанных с устойчи
востью плодов к R. nigricans: сокращалась заражаемость |
плодов |
грибом на 15%, удлинялся инкубационный период развития гри |
|
ба на 10—15 ч и уменьшалась скорость его распространения |
|
па 25—35%- Еще более четкие различия наблюдались |
при ис |
кусственном заражении грибом F. lycopersici Sacc. томатов, об лученных дозой 300 крад при мощностях дозы 28 и 1100 рад/с.
Однако необходимо отметить, что решающую роль в ослабле нии устойчивости плодов играет величина дозы облучения.
Физиологическое состояние плодов. Известно, что у плодов в процессе онтогенеза меняется устойчивость к микроорганизмам. 37-
Так, персики, абрикосы и яблоки при дозревагиги теряют способ ность сопротивляться внедрению патогена.
Значительная потеря устойчивости в результате облучения у зеленых плодов проявляется как в увеличении скорости рас пространения грибов в их тканях и сокращении продолжитель ности инкубационного периода развития гриба, так и в повыше нии их заражаемости. В зрелых облученных плодах таких изме нении не обнаружено (табл. 6).
|
|
|
|
|
Т Л Б Л И Ц Л б |
||
Физиологическое |
|
|
Количество зара |
Продолжитель |
Скорость |
||
|
|
зившихся |
ность инкуба |
распростране |
|||
состояние |
плодов |
Доза |
облуче |
плодов через |
|||
ционного перио |
ния гриба на |
||||||
к моменту |
облуче |
ния, |
крад |
2 дня после |
|||
да развития гри |
поверхности |
||||||
ния |
|
|
|
нпокулирования |
|||
|
|
|
ба, ч |
плодов, мм/ч |
|||
|
|
|
|
гриба, % |
|||
|
|
|
|
|
|
||
|
|
Я б л о к и с о р т а А н т о н о в к а о б ы к н о в е н н а я |
|
||||
Н е в ы з р е в ш н е |
О ( к о н т р о л ь ) |
20,8 |
200 |
0,09 |
|||
|
|
300 |
|
06,5 |
66 |
0,25 |
|
З р е л ы е |
|
0 ( к о н т р о л ь ) |
64,0 |
34 |
0,46 |
||
|
|
300 |
|
91,0 |
28 |
0,51 |
|
|
|
|
П е р с п к н с о р т а Э л ь б е р т а |
|
|||
Н е в ы з р е в ш н е |
0 ( к о н т р о л ь ) |
54,3 |
60 |
0,30 |
|||
|
|
300 |
|
85,0 |
42 |
0,52 |
|
З р е л ы е |
|
0 ( к о н т р о л ь ) |
90,0 |
39 |
0,66 |
||
|
|
300 |
|
100,0 |
38 |
0,74 |
|
|
|
|
А б р и к о с ы с о р т а К р а с н о щ е к и й |
|
|||
Н е в ы з р е в ш н е |
0 ( к о н т р о л ь ) |
50,0 |
45 |
0,31 |
|||
|
|
300 |
|
87,5 |
20 |
0,43 |
|
З р е л ы е |
|
0 ( к о н т р о л ь ) |
100,0 |
23 |
0,71 |
||
|
|
300 |
|
100,0 |
22 |
0,78 |
|
Различная реакция на облучение у плодов разной стадии зрелости отмечена и в опытах с томатами. Кривые, приведенные на рис. 9, показывают, что по мере дозревания заметно ослабля ется сопротивляемость томатов инфекции: чем спелее плоды, тем 38 быстрее распространяется в них гриб. В то же время в процессе

перечисленными выше факторами. У яблок снижение устойчи вости к микроорганизмам всегда более резко выражено в ранние сроки после облучения. По мере хранения различия между конт рольными и облученными плодами становятся менее выражен ными, а различия по дозам практически исчезают.
Из полученных данных следует, что радуризацпп желательно подвергать зрелые плоды. У них резко снижается ипфекцпониая нагрузка и пе происходит столь значительных изменении в при родной устойчивости тканей, как в незрелых плодах.
Сроки внесения инфекции после облучения. В табл. 7 приве дены данные опытов по внесению иифекцпп после облу чения.
Из данных табл. 7 впдпо, что значительные изменения в пло дах происходят при заражении их через сутки после облучения. Через 5 дней существенной разницы в скорости распространения плесневых грибов на облученных и необлучеппых плодах не от мечено. Это можно объяснить как возможной реактивацией растительных объектов от повреждающего действия радиации, так и некоторым спижеппем природной сопротивляемости конт рольных плодов в результате нх более быстрого созревания. Особенпо наглядно это видно в опытах с томатами. При внссеппи инфекции до облучения, как это чаще всего бывает в практике хранения, развитие ее па контрольных н облучепиых плодах происходило одинаково. В даииом случае часть микрофлоры была подавлена действием радиации. Если заражение осущест вляли вскоре после облучения (сразу пли спустя 18—20 ч), об лученные плоды заражались иампого быстрее иеоблучснных, так как сильные культуры грибов вносили в плоды, сопротивляе мость которых подавлена. Прп заражении томатов через 14 сут после облучения разницы в скорости заражения не было, по скольку плоды уже созрели в этот период, причем контрольные быстрее облученных (рис. 10).
Полученные данные имеют большое практическое значение, так как показывают, что отрицательное влияние облучения — снижение природной сопротивляемости — может сопровождать ся положительным фактором — резким снижением инфекцион ной нагрузки. Следовательно, если к моменту восстановления жизнедеятельности спонтанной микрофлоры или развития по павших из воздуха и внедрившихся на плодах микроорганизмов естественная устойчивость «хозяина» — плода — в какой-то мере восстановится, то можно не ожидать быстрого развития «пара зита» на нем.
|
|
|
|
Т А Б Л И Ц А 7 |
|
|
Время (в ч), необходимое |
для образования |
|
|
|
пораженного участка диаметром 10 мм |
||
|
Доза облучения, |
на плодах, |
заражеппых после |
|
Плоды |
облучения |
|
|
|
крад |
|
|
|
|
|
|
|
|
|
|
|
через 1 сут |
через э сут |
через 8 сут |
|
R h i z o p u s |
nigrican s |
|
|
А б р и к о с ы |
0 |
33 |
29 |
20 |
|
300 |
21 |
24 |
20 |
|
|
|
|
|
П е р с и к и |
0 |
49 |
34 |
24 |
|
39 |
30 |
24 |
|
|
300 |
|
|
|
|
P e n i c i l l i u m |
expansum |
|
|
А б р и к о с ы |
0 |
34 |
38 |
28 |
|
300 |
28 |
32 |
22 |
П е р с и к и |
о |
90 |
72 |
45 |
|
300 |
72 |
68 |
42 |
|
M o n i l i a f r u c t i g e n a |
|
|
|
А б р и к о с ы |
0 |
24 |
21 |
16 |
|
300 |
24 |
22 |
12 |
П е р с и к и |
о |
36 |
30 |
24 |
|
300 |
36 |
28 |
24 |
Виды грибов. По отношению к факультативным |
паразитам, |
|||
какими являются плесневые и дрожжевые грибы для плодов, нет полностью иммунных видов или сортов растительного сырья. Есть виды и сорта, лишь относительно устойчивые к определен ным патогенам. Причем устойчивость проявляется в удлинении
инкубационного периода и замедлении скорости |
образования |
гнили, но никак не связана с непоражаемостыо |
растительных |
тканей грибами. |
41 |