

Диабет и мутации митохондрий.
Механизм регуляции высвобождения инсулина β клетками поджелудочной железы тесно связан с уровнем АТФ.
Дефект гена глюкокиназы (изофермент глюкокиназа4) приводит к появлению диабета с фенотипом МОД2. Из-за низкой активности глюкокиназа блокируется секреция инсулина.
Мутация генов тРНК также нарушает выработку АТФ и может возникнуть диабет
II типа.
При генетических мутациях НАДФ трансгидрогеназы-важного механизма защиты от АФК, повреждение митохондрий уменьшает выработку АТФ, блокируя высвобождение инсулина β клетками.
Итак, мутации митохондриального генома накапливаются в течение жизни организма. Мутации в генах, которые кодируют компоненты дыхательной цепи, АТФсинтазу, систему защиты от АФК, а также мутации в генах тРНК вызывают у
человека ряд заболеваний, чаще всего поражающих мышечный аппарат, сердце, мозг, поджелудочную железу.
Полиморфизм гена апобелка Е, клиническое значение.
Аллельный полиморфизм гена aполипопротеина Е (ApoE) - генетический маркер риска нарушений липидного обмена
Исследование аллельного полиморфизма гена ApoE имеет прогностическое значение, позволяющее определить риск развития ишемической болезни сердца вследствие нарушения липидного обмена.
Ген ApoE кодирует аминокислотную последовательность белка aполипопротеина Е. Aполипопротеин Е синтезируется в печени и головном мозге, и играет существенную роль в метаболизме липидов. Aполипопротеин Е входит в состав жировых частиц хиломикронов и липопротеинов очень низкой плотности (ЛПОНП), инициируя их захват и удаление из крови через взаимодействие со специфическим рецептором на поверхности клеток печени. АроЕ участвует в некоторых других процессах, таких как иммунорегуляция, нервная регенерация и активация некоторых липолитических ферментов (липазы печени, липазы липопротеинов и лецитин-холестерин ацилтрансферазы). Он необходим для доставки холестерина от глиальных клеток мозга до нейронов. Эффективность взаимодействия ApoE с рецепторами определяется уникальным строением белковой молекулы.
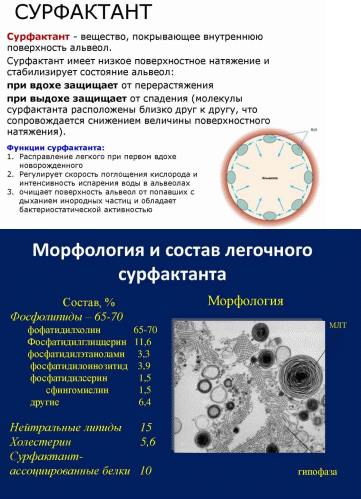
Биохимические основы синдрома дыхательных расстройств. Строение сурфактатнта.

Лабораторная диагностика панкреатитов.

Молекулярные механизмы патогенеза острого панкреатита.

Патогенетические особенности нейродегенеративных заболеваний нервной системы.
Амилоидоз - маркер нейродегенеративных заболеваний.
Такие заболевания как болезнь Альцгеймера, Хантингтона,
Паркенсона являются результатом нарушения нормального процесса формирования структуры белка. Растворимый белок за счет неправильного фолдинга белка превращается в нерастворимые внеклеточные амилодные волокна. Такие заболевания относят к амилоидозам.
Отложение амилоида приводит к разрушению клеток. Болезнь Альцгеймера вызвана отложением внеклеточного амилоида нейронами при участии белка βамилоида. Отложение амилоида является одной из ведущих причин заболевания, кроме того у таких больных происходит агрегация белка тау, который вызывает лобновисочную деменцию. С возрастом значительно увеличивается количество окисленных белков, в ходе таких реакций образуются альдегиды и свободные радикалы. Образующиеся β-амилоидные агрегты усиливают образование пероксида водорода, формируется окислительный стресс.
Сывороточный белок амилоид яляется нормальным белком- предшественником фибрилярного тканевого белка, синтезирующегося в печени.
Амилоид А является маркером острой фазы воспаления.
Стимулятором его увеличения в сыворотке является интерлейкин1. Амилоид явялется фибрилярным белком, амилоидоз-диспротеиноз сопровождается глубоким нарушение белкового обмена, появлением в межуточной ткани и стенках сосудов амилоида, то есть – амилоидных фибрилл, которая у людей с болезнью Альцгеймера откладывается в виде бляшек на поверхности нейронов. В амилоиде много β слоев, ориентированных перепендикулярно амилоидной фибрилле. В β амилоидном белке возникают вытянутые двухслойные параллеьные β
складки, которые могут взаимодействовать друг с другом, образовывая волокна. Прогрессирующее накопление бляшек β амилоида связанны с нарушением 3-х генов: 1) с мутациями в гене его предшественника; 2) мутации в генах предшественников
прессенилинов (PS-1, PS-2), что способствует секреции β амилоида и образованию его формы с повышенной склоностью к агрегации. Амилоидозы бывают вызваны наследуемыми мутациями лизоцима, αцепи фибриногена А, аполипотеинов А1, АII. Амилоидный белок обычно секретируется только пораженой тканью, так что его высокая концентрация вокруг этой ткани приводит к отложению амилоидных волокон.
Амилоидозы относятся к системным заболеваниям. Первичный системный амилоидоз может быть вызван отложением волокон, состоящих из неправильно свернутых легких цепей иммуноглобулинов или их фрагментов.
У пациентов с хроническими ифекционными или воспалительными заболеваниями, такими как ревматоидный артрит, туберкулез, кистозный фиброз, некоторые формы рака усиливается секреция амилоида А. Этот белок в виде волокон откладывается в селезенке, почках, печени, в области сердца- это вторичный амилоидоз. β-амилоид, который играет главную роль в возникновении болезни Альцгеймера происходит из большого тансмембранного белка, который называется белком-предшественником β-амилоида или АРР ( amiloid-β precursor protein), он обнаружен во многих тканях.
38. Маркеры заболеваний бронхолёгочной системы. Механизмы повреждающего действия полимеров α1-антитрипсина.
α-1антитрипсин относится к белкам острой фазы. Воспалительные процессы могут усиливать его образование и способствовать повреждению печени.
Основная функция α-1антиприпсина -защита легких от действия эластазы нейтрофилов. Выраженный дефицит ингибитора ведет к нарушению физиологического баланса между протеазами и ингибиторами в ткани легких, следствием чего является раннее развитие энфиземы легких, и респираторного дистресс-синдрома.
α -1антитрипсин является гликопротеидом, синтезируемым в печени. Он обеспечивает
90% активности, ингибирующей трипсин в крови, α-1антитрипсин тормозит не только действие трипсина, но и химотрипсина, эластазы, калликреина, катепсинов, и других тканевых протеаз.
Ингибиторы сериновых протеаз (серпины) – суперсемейство белков, широко распространенных в живых организмах, включая растения, вирусы, животных. К ним относятся α -1антитрипсин, ингибитор фактора С1- комплемента, антитромбин III, ингибитор активатора плазминогена 1 (PAJ 1) занимающих ключевые позиции в процессах воспаления, системе комплемента, коагуляции и фибринолизе.
Все члены суперсемейства серпинов имеют с α -1 антитрипсином схожесть третичной структуры.
В основе пространственной структуры серпинов лежат 3 участка, уложенных в форме β структуры (А-С) и подвижная петля, которая представляет последовательность аминокислот, играющих роль псевдосубстрата для протеазы. У α -1 антитрипсина в состав петли входит последовательность метионин-серин, выполняющий функцию «приманки» для эластазы нейирофилов. Эластаза, связавшись с ингибитором при участии петли, неспособна катализировать гидролиз пептидной связи. Этот комплекс узнается печеночными рецепторами и удаляется из кровотока.
Динамика содержания α -1антитрипсина при воспалительных процессах.
Содержание α -1антитрипсина повышается при воспалительных процессах:
Острых и хронических инфекционных заболеваний;
Гепатитах;
При вакцинации;
Послеоперационных состояниях; Остром и хроническом панкреатите; При злокачественных образованиях.
Понижается:
Врожденный дефицит ингибитора, сочетающийся с ювенильной базальной энфиземой легких, муковисцидозом;
При вирусном гепатите (нарушение синтеза в печени);
Респираторный дистресс-синдром;
Коагулопатия;
В острой фазе термической травмы.
