

7. Окислительное фосфорилирование: сущность, биологическое значение процесса.
Энергия, образующаяся при прохождении потока электронов по дыхательной цепи, используется для сопряженного фосфорилирования АДФ. Эти два
процесса взаимозависимы: окисление не может протекать в отсутствие АДФ.
Соотношение окисления и фосфорилирования определяется коэффициентом
P/O, показывающим, какое количество Рi используется на образование АТФ
при превращении одного грамм-атома О2 в Н2О (количество моль фосфорилированного АДФ на 1/2 моль кислорода). Коэффициент Р/О называется коэффициентом окислительного фосфорилирования и зависит от точки вхождения восстановительных эквивалентов в цепь транспорта электронов. Например, для субстратов, окисляемых NAD-зависимой дегидрогеназой, Р/О=3, так как в дыхательной цепи есть три участка, где перенос электронов сопряжен с синтезом АTФ. Не все субстраты передают электроны и протоны на NAD, некоторые окисляются FAD-зависимыми дегидрогеназами, которые переносят протоны иэлектроны сразу на убихинон, минуя комплекс I. В этом случае Р/О=2. В действительности коэффициент фосфорилирования всегда меньше теоретической
величины, потому что часть энергии, высвобождающейся при транспорте электронов, расходуется не на синтез АТФ, а для переноса веществ через митохондриальную мембрану.
В сутки человек потребляет в среднем 27 моль кислорода. Основное его количество (примерно 25 моль) используется в митохондриях в дыхательной цепи.
Следовательно, ежесуточно синтезируется 125 моль АTФ или 62 кг (при расчетеиспользовали коэффициент Р/О=2,5, то есть среднее значение коэффициентафосфорилирования). Масса всей АТФ, содержащейся в организме, составляетпримерно 20–30 г. Итак, можно сделать вывод, что каждая молекула АTФ за сутки 2500 раз проходит процесс гидролиза и синтеза, что и характеризует интенсивность обмена АTФ.
8. Цепь переноса электронов (ЦПЭ), сопряжение дыхания и
синтеза АТФ в митохондриях, коэффициент окислительного
фосфорилирования. Ингибиторы и разобщители ЦПЭ.
Цепь переноса электронов (ЦПЭ)
Имеется два пути синтеза АТФ в клетке: 1) окислительное фосфорилирование и 2) субстратное фосфорилирование.
Окислительное фосфорилирование является главным путем синтеза АТФ из АДФ. С помощью этого механизма в клетках миокарда и скелетных мышц синтезируется около 90% АТФ. Реакция энергетически сопряжена с переносом электронов с восстановленных коферментов на кислород и требует наличия неповрежденной митохондриальной мембраны. Альтернативный путь синтеза АТФ из АДФ – субстратное фосфорилирование. В этом случае происходит перенос фосфата с высокоэнергетических фосфатов клетки на АДФ с образованием АТФ. Это происходит в ходе креатинкиназной реакции:
а) Креатинфосфат + АДФ → креатин + АТФ
и в ходе 2-х реакций гликолиза: фосфоглицераткиназной и пируваткиназной
б) 1,3-дифосфоглицерат + АДФ → 3-фосфоглицерат + АТФ
в) фосфоенолпируват + АДФ → пируват + АТФ
В этом случае механизм сопряжения не нуждается в наличии кислорода и неповрежденной митохондриальной мембраны.
Дыхательная цепь
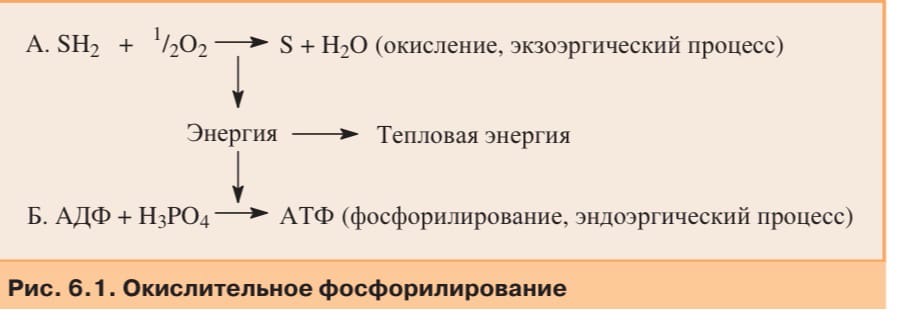
Окисление субстратов в процессе дыхания можно представить как перенос электронов и протонов от органических веществ на кислород:
SH2+ ½O2®S+H2O
Перенос электронов на кислород происходит при участии системы переносчиков, встроенных во внутреннюю мембрану митохондрий и образующих цепь переноса
электронов (ЦПЭ) или дыхательную цепь. В состав ЦПЭ входят 4 ферментативных комплекса: НАДН-дегидрогеназа (I), сукцинатдегидрогеназа (II),Q·H2-дегидрогеназа, цитохромоксидаза (IV), а также 2 низкомолекулярных переносчика: гидрофобная молекула коферментаQи цитохром С – небольшой по размерам белок. Все компоненты ЦПЭ расположены в митохондриальной мембране в порядке возрастания редокс потенциала (окислительно-восстановительного потенциала); самый высокий редокс–потенциал у кислорода. Это обеспечивает последовательное перемещение электронов от НАДН на кислород, при этом происходит выделение энергии на каждом этапе ЦПЭ.

Рис. 1. Митохондриальная цепь переноса электронов.
Атомы водорода или электроны перемещаются по цепи от более электроотрицательных компонентов к более электроположительному кислороду. Изменение окислительно-восстановительного потенциала при переходе от системы НАД+/НАДН к системе О2/Н2О составляет 1,1 В.
Водород от первичных доноров вводится в дыхательную цепь с участием НАД-зависимых (цитрат, 2-оксоглутарат, малат, пируват, глутамат) или ФАД-зависимых дегидрогеназ (сукцинат, ацил-КоА, глицерол-3-фосфат). НАД-зависимые дегидрогеназы переносят водород на НАД+(с образованием НАДН·Н+), а ФАД-зависимые дегидрогеназы – на ФАД (с образованием ФАД·Н2). Далее с НАДН·Н+водород при участии НАДН-дегидрогеназы (Iкомплекса) передается на КоQс образованием КоQ·H2. НАДН-ДГ представляет собой ФМН-содержащий фермент. С ФАД·Н2водород при участии СДГ (IIкомплекс) также передается на КоQс образованием КоQ·H2. КоQили убихинон является компонентом митохондриальных липидов. Его содержание значительно превосходит содержание других компонентов дыхательной цепи, поэтому считается, что КоQявляется подвижным компонентом дыхательной цепи, который получает восстановительные эквиваленты от фиксированных флавопротеиновых комплексов и передает их на цитохромы. Цитохромы – это переносящие электроны белки, молекула которых содержит в качестве простетической группы гем. Цитохромы дыхательной цепи выстроены в порядке возрастания окислительно-восстановительного потенциала и обозначаются латинскими буквами:b,c1,c,aиa3. Терминальный цитохром а-а3, называемый также цитохромоксидазой, осуществляет перенос восстановительных эквивалентов на молекулярный кислород. Цитохромоксидаза содержит медь и имеет очень высокое сродство к кислороду, что позволяет дыхательной цепи функционировать с максимальной скоростью до тех пор, пока в ткани не будет практически исчерпан кислород:
О2+ 4e+ 4H+®2 Н2О
Дополнительным компонентом, находящимся в дыхательной цепи, является железо-серный белок, FeS(негемовое железо). Полагают, что он участвует в окислительно-восстановительном процессе, протекающем по одноэлектронному механизму.
Для объяснения механизма сопряжения окисления и фосфорилирования было выдвинуто несколько гипотез, из которых экспериментальное подтверждение получила только хемиосмотическая теория, предложенная Питером Митчеллом в 1961 году. Согласно этой теории первичным процессом в окислительном фосфорилировании является транслокация протонов (Н+) на наружную сторону внутренней митохондриальной мембраны. При этом предполагается, что мембрана непроницаема для ионов вообще и особенно для протонов, которые накапливаются на наружной стороне мембраны, создавая по обе стороны мембраны разность электрохимических потенциалов (Dmн+). Она складывается из химического потенциала (разность рН) и электрического потенциала. Разность электрохимических потенциалов обеспечивает действие локализованной в мембране АТФ-синтазы, которая в присутствии АДФ и Фн синтезирует АТФ.
Предполагается, что дыхательная цепь уложена в мембране в виде трех окислительно-восстановительных петель, каждая из которых состоит из переносчиков водорода и переносчиков электронов:
АТФ-синтаза по своей структуре похожа на гриб. При этом шляпка гриба, представляющая собой фосфорилирующие субъединицы (F1-субъединица), ответственные за образование АТФ, находятся на поверхности внутренней мембраны, обращенной в матрикс. А ножка гриба представляет собой канальную часть (Fo-субъединица) АТФ-синтазы, пронизывающей всю толщу внутренней мембраны. Электрохимический потенциал заставляет протоны двигаться с наружной поверхности внутрь, кF1-субъединице, обладающей каталитической активностью, т.е. способностью катализировать образование АТФ из АДФ и Фн (Н3РО4).
Коэффициент фосфорилирования
Оценка эффективности улавливания энергии интактными митохондриями показывает, что при окислении субстратов, идущем с участием НАД-зависимых дегидрогеназ и дыхательной цепи, происходит включение 3 молей неорганического фосфата в АДФ и образуется 3 моля АТФ на ½ моля потребленного кислорода. Отношение Р/О (коэффициент фосфорилирования) = 3. В то же время при окислении субстрата через флавопротеиновую дегидрогеназу образуется только 2 моля АТФ, т.е. Р/О = 2.

Ингибиторы тканевого дыхания
Ингибиторы окисления прерывают поток электронов на участках дыхательной цепи, блокируя одно из трех звеньев образования протонного потенциала, Первая группа препаратов блокирует поток электронов на участке между НАД и КоQ. К этим препаратам относятся барбитураты - амитал или барбамил (малые транквилизаторы), ротенон (инсектицид), пиерицидин (антибиотик).
Вторая группа ингибиторов окисления, к которым относится антимицин А (антибиотик) блокирует поток электронов на участке цит. b566-FeS-белок, т.е. на уровне второго звена сопряжения.
Третья группа ингибиторов окисления, к которым относятся цианиды, азиды, оксид углерода, сероводород блокируют цитохромоксидазу.
Ингибиторы фосфорилирования действуют на АТФ-синтазу, препятствуя использованию протонного потенциала для синтеза АТФ. К этим веществам относятся олигомицин и дициклокарбодиимид.
УЧЕБНИК
Цепь переноса электронов — ЦПЭ
Указанное выше уравнение для окислительно-восстановительной реакции
представляет собой обобщенную форму, так как изображает процесс окисления субстратов как прямое дегидрирование, причем кислород выступает в роли непосредственного акцептора водорода. На самом деле кислород получает электроны иным образом. Существуют промежуточные переносчики при транспорте электронов от исходного донора электронов SH2 к терминальному акцептору — О2. Полный процесс представляет собой цепь последовательных окислительно-восстановительных реакций, в ходе которых происходит взаимодействие между переносчиками. Каждый промежуточный переносчик вначале выступает в роли акцептора электронов и протонов и из окисленного состояния переходит в восстановленную форму. Затем он передает электрон следующему переносчику и снова возвращается в окисленное состояние. На последней стадии переносчик передает
электроны кислороду, который затем восстанавливается до воды. Совокупность последовательных окислительно-восстановительных реакций называется цепью переноса (транспорта) электронов, или дыхательной цепью (рис. 6.2).

Промежуточными переносчиками в дыхательной цепи у высших организмов
являются коферменты: NAD+ (никотинамид-адениндинуклеотид), FAD и FMN(флавинадениндинуклеотид и флавинмононуклеотид), кофермент Q (CoQ), семейство гемсодержащих белков — цитохромов (обозначаемых как цитохромы b,c1, c, a, a3) и белки, содержащие негеминовое железо. Все участники этой цепи организованы в четыре окислительно-восстановительных комплекса (рис. 6.5), связанные убихиноном (CoQ) и цитохромом c.
Процесс начинается с переноса протонов и электронов от окисляемого
субстрата на коферменты NAD+ или FAD. Это определяется тем, является ли
дегидрогеназа, катализирующая первую стадию, NAD-зависимой или FADзависимой. NAD-зависимая дегидрогеназа катализирует реакции окисления непосредственно субстрата (первичная дегидрогеназа). NAD+ является коферментом и выполняет роль акцептора водорода (рис. 6.3). FAD-зависимая дегидрогеназа также выполняет функцию первичной дегидрогеназы. Кофермент FAD является акцептором водорода от субстрата. Если процесс начинается с NAD+, то следующим переносчиком будет NADH-дегидрогеназа, коферментом которой является FMN.

Тип участвующей дегидрогеназы зависит от природы субстрата. Но каким бы
ни был исходный субстрат, электроны и протоны от флавинов переносятся к
коферменту Q, а дальше пути электронов и протонов расходятся. Электроны с помощью системы цитохромов достигают кислорода, который затем, присоединяя протоны, превращается в воду. Чтобы разобраться в системе транспорта электронов, необходимо познакомиться с отдельными ее участниками.
Символ 2Н+ означает два протона, обычно переносимые в виде гидрида
иона. В этом случае вместо терминов «донор электронов» и «акцептор электронов» иногда используют термины «донор или акцептор водорода».
NADH-дегидрогеназа катализирует окисление NADH и восстановление
убихинона (CoQ). Переносчиком водорода является кофермент — FMN (комплекс I на рис. 6.5). Строение FMN представлено на рис. 6.3. В процессе реакции водород сначала присоединяется к FMN, соединенному с ферментом, а затем протоны поступают в межмембранное пространство, а электроны с помощью FeS-белков передаются на убихинон. Флавиновые коферменты (FAD и FMN) содержат производные витамина B2 и прочно связаны с ферментом, поэтому ферменты, в состав которых они входят, называются флавопротеинами. Акцептором электронов от комплекса I является убихинон (кофермент Q) — производное изопрена, его название возникло из-за повсеместной распространенности в природе.
Получая 2e из матрикса митохондрий, он восстанавливается в убихинол. Кофермент Q действует как переносчик электронов на цитохромы. Цитохромы — это гемопротеины, содержащие в качестве простетической группы гем, прочно связанный с белковой частью. Атом железа в геме может менять валентность, присоединяя или отдавая электроны:
В дыхательной цепи цитохромы служат переносчиками электронов и располагаются соответственно величине окислительно-восстановительного потенциала следующим образом: b, c1, c, а, а3. Гемовые группы цитохромов связаны сбелковой частью координационными связями между ионом железа и соответствующими аминокислотными остатками.
В цитохромах c и c1 дополнительные ковалентные связи формируются между тиоловыми группами цистеина и боковыми винильными группами гема. QН2-дегидрогеназа (комплекс III на рис. 6.5) представляет собой комплекс цитохромов b и c1. Этот фермент катализирует окисление восстановленногокофермента Q и перенос электронов на цитохром c. Электроны последовательно переносятся атомами железа цитохромов b и c1, а затем поступают на цитохром c. В то же время протоны за счет энергии, выделяющейся в процессе транспорта e- с цитохрома b на цитохром с, выделяются в межмембранное пространство. Цитохромоксидаза включает комплекс цитохромов а и а3 (комплекс IV на рис. 6.5). Цитохромоксидаза кроме гема содержит ионы меди, которые способны менять валентность и таким способом участвовать в переносе электронов:
Цитохромоксидаза переносит электроны с цитохрома c на кислород. В переносе электронов участвуют сначала ионы железа цитохромов а и а3, а затем ион меди цитохрома а3. Молекула кислорода связывается с железом в геме цитохрома а3. Следовательно, переход электронов на кислород с иона меди цитохромаа3 происходит на молекуле фермента. Каждый из атомов молекулы кислородаприсоединяет по два электрона и протона, образуя при этом молекулу воды.
