

Неотложные состояния в педиатрии. Сидельников В.М
..pdfросекреция АДГ. Повышение уровня АДГ оказывает влияние на гиалуронидазу. Последняя изменяет «пористость» мембраны почечных канальцев, увеличивая проницаемость почечных канальцев для воды, т. е. повышается реабсорбция воды и уменьшается ее выведение из организма. Гипонатриемия вызывает уменьшение выработки АДГ, и, следовательно, повышается выведение жидкости из организма. Одновременно с ответной реакцией осморецепторов у человека появляются ощущение жажды, сухости слизистой оболочки полости рта и желание пополнить организм водой. У ребенка это ощущение сопровождается плачем и криком. Считают, что эти импульсы поступают из осморецепторов к коре большого мозга, так как ощущение жажды дифференцируется в сознании даже у новорожденных.
Таким образом, при гипернатриемии у человека появляется жажда, увеличивается потребление воды и уменьшается диурез за счет повышения содержания АДГ. Гипонатриемия вызывает обратное: ощущение жажды уменьшается, а затем исчезает, диурез увеличивается, потребление жидкости снижается.
Описанные процессы продолжаются, пока не установится физиологическая концентрация натрия. В процессе обратной связи осмотическая регуляция обеспечивает поддержание на нормальном уровне содержания натрия.
На изменение объема внеклеточной жидкости реагируют рецепторы, находящиеся в почечных клубочках (юкстагломерулярного аппарата). Точнее, эти рецепторы реагируют не на изменение объема жидкости, а на его гемодинамические последствия, т. е. на изменение кровотока.
В ответ на ухудшение кровоснабжения почек усиливается выработка ренина вюкстагломерулярном аппарате, который преобразуется в ангиотензиноген (аг-глобулин), а в дальнейшем в ангиотензин, вызывающий вазопрессорный эффект и стимулирующий выработку альдостерона надпочечниками. Альдостерон увеличивает процессы реабсорбции натрия.
При улучшении кровообращения уменьшается выработка ренина, что в конечном итоге уменьшает выработку альдостерона. В ответ выведение натрия почками увеличивается.
Регуляция выделения и реабсорбции воды и натрия зависит в значительной мере от общего ОЦК и степени возбуждения волюморецепторов. Существование волюморецепторов доказано для левого и правого предсердий, для устьев легочных вен и некоторых артериальных стволов.
Импульсы от волюморецепторов левого предсердия поступают в ядра гипоталамуса и влияют на секрецию АДГ. При увеличении ОЦК выработка АДГ снижается, а при уменьшении ОЦК — повышается.
21
Импульсы от волюморецепторов правого предсердия поступают в центры, расположенные в задней части гипофиза и передней части среднего мозга, связанные с эпифизом. Раздражение центров стимулирует образование адреногломерулотропица, который активирует выработку альдостерона. Усиленный синтез альдостерона увеличивает реабсорбцию натрия в почечных канальцах.
Важное значение в регуляции водно-электролитного обмена имеют внепочечные механизмы: кожа, органы дыхания, пищеварение, печень, селезенка, различные отделы ЦНС и эндокринные железы.
До сих пор остается нераскрытой проблема так называемого солевого выбора при недостаточном поступлении в организм тех или иных элементов. В таких случаях животные предпочитают пищу, содержащую эти недостающие элементы, и наоборот, при избыточном поступлении в организм элемента отмечается снижение аппетита к пище, содержащей его. По-видимому, в этих случаях важную роль играют специфические рецепторы внутренних органов.
2. КИСЛОТНО-ОСНОВНОЕ СОСТОЯНИЕ
ОРГАНИЗМА, ЕГО РЕГУЛЯЦИЯ И ПРИНЦИПЫ
КОРРЕКЦИИ НАРУШЕНИЙ
В процессе жизнедеятельности организма происходит накопление большего количества кислот, чем оснований. Кроме того, в нормальном пищевом рационе человека количество кислых продуктов значительно превышает количество щелочных. Таким образом, в процессе питания и метаболизма в организме создаются условия для закисления различных внутренних сред. В процессе эволюции в человеческом организме выработались регуляторные механизмы, обеспечивающие строгое постоянство реакций внутренней среды. Такими механизмами являются физи- ко-химические системы крови (буферные системы) и физиологические системы регуляции кислотно-основного состояния (КОС), осуществляемые легкими, почками, печенью, пищеварительным каналом. Указанные механизмы позволяют поддерживать рН крови на постоянном уровне, колеблющемся в пределах 7,35— 7,45, т. е. слабо щелочной реакции крови (табл. 9). Колебания рН крови в этих пределах необходимы в связи с тем, что при дан-
Таблица 9 Колебания показателей КОС в человеческом организме
6,8 |
|
7,35 |
7,45 |
7,8 |
|
|
|
|
|
I IccouMecTUMO |
с |
Ацидоз |
Алкалоз |
Несовместимо с |
жизнью |
|
|
|
жизнью |
22

.них показателях обеспечивается оптимальная активность ферментов человеческих сред. При отклонениях рН крови как в кислую, так и в щелочную сторону прекращается деятельность ферментативных систем. При далеко зашедших сдвигах рН крови в человеческом организме возникает ситуация несовместимости с жизнью, когда клетки организма не в состоянии поддерживать свою жизнедеятельность. Таким образом, постоянство рН внутренней среды — изогидрия — является важнейшей константой жизни организма.
В норме в организме взрослого человека ежедневно образуется 40—80 ммоль Η ' и 1300 ммоль СО2. Углекислота через свою гидрированную форму — угольную кислоту — становится источником Η>":
, причем Н2СОз обра-
зуется со скоростью 10 мэкв/мин.
У здорового человека баланс метаболических Н+ равен «0»,
т.е. количество образованных Н+ равно количеству выводимых. Сохранение гомеостаза рН биологических жидкостей осу-
ществляется следующими регуляторными механизмами:
1. Механизмы разведения и метаболирования Н+ в общем объеме жидкостей организма.
2.Буферные, или физико-химические, системы организма.
3.Физиологические системы регуляции КОС.
Механизмы разведения и метаболирования Η '" — простейший механизм регуляции КОС, предупреждающий резкие отклонения рН внеклеточной жидкости.
Образовавшиеся кислоты в организме подвергаются следующим изменениям:
1.Одна часть их окисляется в клетках до СОг и НгО.
2.Другая часть (нелетучие кислоты) вступает в различные обменные процессы, и эти вещества перестают существовать как кислоты.
3.Третья часть переходит из внутриклеточных пространств во внеклеточное и выводится с мочой.
Буферные системы организма. Буферными растворами называют такие растворы, рН которых не изменяется при добавлении оснований и кислот. К буферным системам организма относятся: бикарбонатная (угольная кислота), гемоглобиновая, протеиновая, система плазменных белков, фосфатная (система неорганический двунатриевый — мононатриевый фосфат и система органических фосфатов).
Количественное обеспечение буферных емкостей в поддержании КОС распределяется следующим образом: гемоглобиновая буферная система — 73—76 %, бикарбонатная буферная система 12—27 %, протеиновая система — 2- -5 %, фосфатная буферная система — 1—2 %.
23

Бикарбонатная буферная система (угольная кислота).
В результате окислительных процессов в организме постоянно образуется диоксид углерода (ССЬ), который в водной среде превращается в угольную кислоту. Последняя диссоциирует на ионы Н+ и НСОз~, образующие соединения с веществами плазмы, преимущественно с Na+. Таким образом, в крови одновременно существует угольная кислота и ее соли. Между ними устанавливается строго постоянное взаимодействие, важнейшим условием которого является постоянное соотношение этих веществ, или постоянство константы, выражающееся соотношением 20 : 1 независимо от общего их содержания. Эти процессы можно изобразить в виде следующих химических реакций:
Снижение этого соотношения ведет к ацидозу, а повыше-
ние — к алкалозу. |
|
Между Н + , |
и Н2СО3 существует строгая зависимость, |
которую определил Henderson формулой
Hasselbach эту величину привел в отрицательную логарифмическую форму:
где рК. представляет отрицательный логарифм диссоциации угольной кислоты, который равен 6,1 при стандартных условиях.
Зная, что в норме в крови-содержится 27 мэкв/л бикарбонатов и 1,35 мэкв/л угольной кислоты, можно по уравнению Henderson— Hasselbach определить нормальные величины рН крови, т. е.
Образование кислых продуктов в организме нейтрализуется бикарбонатами плазмы крови с образованием эквивалентного количества слабой угольной кислоты. Избыток последней диссоциирует на воду и диоксид углерода. Выведение диоксида углерода осуществляется легкими, что обусловливает восстановление нарушенного равновесия, бикарбонат / угольная кислота, благодаря чему сохраняется постоянство рН.
Таким образом, согласно уравнению Henderson—Hassel-
24

bach, реакция рН крови зависит не от абсолютных количеств бикарбоната и угольной кислоты, а от их соотношения. Реакция рН крови остается неизмененной при одинаковых изменениях числителя и знаменателя, рН крови изменяется, если изменяется только числитель или только знаменатель.
Фосфатная буферная система крови. Сущность этой буферной системы заключается в том, что однозамещенный фосфат (Nal-bPCXi), обладающий слабо кислыми свойствами, превращается а двузамещенный фосфат (Na2HPO4), которому присущи основные свойства. Фосфатная буферная система является главной буферной системой клеток и мочи. Механизм компенсации сводится к образованию подвижных фосфатов в клетке и фосфатных солей мочи.
Протеиновая буферная система крови. Буферные свойства белков крови обусловлены тем, что белки являются амфотерными электролитами, или амфолитами. Формулу белков можно схематично представить как:
В основном растворе белки отщепляют Н+ и ведут себя как кислота, а в кислом—ОН~ и ведут себя как основания, т. е. белки диссоциируют с образованием как Н+, так и гидроксильных ионов (ОН") в зависимости от того, в каком растворе (основном или кислотном) они находятся.
В норме реакция крови слабо основная (рН — 7,4), в связи с чем белки (в том числе гемоглобин) ведут себя как слабые кислоты.
Гемоглобиновая буферная система является мощной буферной системой. Процессе газообмена в легких и тканях, связывание и отдача СЬ и СОг приводят к постоянным колебаниям КОС, которые, однако, не выходят за пределы физиологических колебаний. При этом значительная роль принадлежит «эффекту Амбурже» — обмену ионов между плазмой крови и эритроцитами (рис. 3). Действие этой системы обусловлено наличием двух форм гемоглобина: восстановленного гемоглобина и оксигемоглобина. Последний обладает более выраженными кислотными свойствами, которые в 80 раз превосходят кислотные свойства восстановленного гемоглобина.
На уровне тканей оксигемоглобин (КНЬОг) расщепляется, при этом кислород поступает в ткани и образуется восстановленный гемоглобин (КНЬ). В свою очередь, углекислота (СО2) поступает в эритроцит и под воздействием карбоангидразы превращается в угольную кислоту, более сильную, чем вос-
становленный гемоглобин. При взаимодействии Н2СОз + КНЬ образуется соединение КНСО3, которое диссоциирует на К+ и НСОг· Анион НСОз" перемещается из эритроцита в плазму и
25
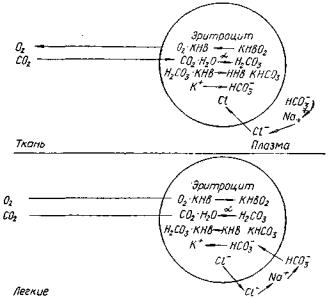
Рис. 3. Роль эритроцитов в поддержании КОС во внеклеточном жидкости (эффект Амбурже)
пополняет ее основные резервы. Одновременно С1~ перемещается в эритроцит, он легко проникает через мембрану последнего.
В легких происходит обратный процесс, так как оксигемоглобин обладает свойствами более сильной кислоты, чем угольная.
Эффект Бора. Парциальное напряжение кислорода в крови зависит от рН. Сдвиг рН в кислую сторону (выход COL> из тканей) способствует диссоциации оксигемоглобина и отдаче кислорода. При повышении рН кровь насыщается меньшим количеством кислорода, кислородная емкость крови возрастает.
Физиологические системы регуляции КОС. М е х а н и з м ы р е г у л я ц и и К О С с у ч а с т и е м л е г к и х . В физиологических условиях дыхательная регуляция осуществляется путем окисления и редуцирования гемоглобина.
Свободные Н5" не выделяются легкими. При усиленном образовании H^ бикарбонат натрия связывает Н+ и переводит сильные кислоты в слабую угольную кислоту с последующим образованием НгО и СО2 (конечные продукты метаболических процессов). СОг является специфическим раздражителем дыхательного центра. Малейшее повышение содержания СО2 вызывает раздражение дыхательного центра, возникает гипервентиляция, и избыток СОг выводится из организма.
При избыточном накоплении основных веществ в организме
26

Канальи, еВоя моча |
Нояальце&ь/е клетки |
Венозная кроЬь & |
шлубочкобый фильтрат) |
|
капиллярах канальцев |
Рис. 4. Схема ацидогенеза
снижается концентрация С02 . В результате интенсивность стимуляции дыхательного центра уменьшается; возникают гиповентиляция м брадипноэ, что приводит к сохранению СО2 и восполнению запасов Н2СОз.
Легкие, по сравнению с почками, выделяют за 24 ч приблизительно в 200 раз больше кислых продуктов в форме летучих кислот (СО2), что постоянно поддерживает на нормальном уровне соотношение бикарбоната и угольной кислоты.
Р о л ь п о ч е к в п о д д е р ж а н и и КОС . Почки путем ацидогенеза и аммониогенеза играют существенную роль в поддержании КОС.
Ацидогенез. Роль почек в сохранении КОС заключается в выведении из кислой крови Η + и из основной — НСОз" (рис. 4). В клетках почечных канальцев под воздействием карбоангидразы происходит реакция
Н2СО3 диссоциирует на Н+ и НСО;Г- В канальцевую мочу поступает бикарбонат натрия, который диссоциирует на Na4 и НСОг. Н+ из клеток почечных канальцев поступают в канальцевую мочу и, соединясь с НСО3 , образуют Н2СО3, из которой диссоциируют Н2О и СО2. В свою очередь, натрий из канальцевой мочи поступает в канальцевые клетки и, соединяясь с НСО(", в виде бикарбоната натрия поступает в венозную кровь капилляров канальцев. СО2, образовавшийся в канальцевой моче, поступает в канальцевые клетки, где также под влиянием карбо-
27

Нокальцевая моча |
Канольцебые клетки |
венозная кроВь |
(клубочкоЬыО срильтрат) |
|
8 капиллярах каноАьцбб |
Рис. 5. Схема аммониогенеза
ангидразы и реакции С02 + Н2 0 образуется угольная кислота НгСОз, которая диссоциирует на Н+ и НСОг·
В канальцевой моче содержится соль фосфата натрия (Na2HPO4). которая диссоциирует на Na+ и НРО?~. Hf из канальцевой клетки перемещается в канальцевую мочу, где образуется NaH2PO4 — кислая соль фосфата натрия, которая выводится с мочой. Na+ из канальцевой мочи перемещается в канальцевые клетки, где соединяется с НСОг и в виде NaHCO3 поступает в венозную кровь капилляров канальцев.
Аммониогснез. В канальцевых клетках под влиянием карбоангидразы из ССЬ и НгО образуется угольная кислота (рис. 5), Последняя диссоциирует на H^ и НСОг- В дальнейшем Н+ переходит в канальцевую мочу. В канальцевой моче NaCI диссоциирует на Na+ и CI". Na f перемещается в канальцевые клетки, где образует соединение с НСОг и в виде бикарбоната проникает в венозную кровь капилляров канальцев.
Кроме того, в клетках почечных канальцев происходит образование аммиака (ЫНз) за счет дезаминирования некоторых аминокислот, в частности из глютаминовой, под влиянием глютаминазы. Аммиак легко диффундирует в канальцевую мочу, где соединяется с Н+, образуя ионы аммония (NHi1"), которые не способны проникать через клеточную мембрану. В канальцах
NH4 ! , соединяясь с С1~", образуют аммония |
хлорид (NH4CI), |
||
который выводится из организма с мочой. |
|
|
|
З н а ч е н и е |
п и щ е в а р и т е л ь н о г о |
к а н а л а |
в |
п о д д е р ж а н и и КОС . В париетальных клетках слизистой оболочки желудка и кишок под влиянием карбоангидразы из ЬЬО и СОг образуется угольная кислота (НгСОз).
Из клеток желудка в его просвет поступает Н+, а НСО3" — в ток крови. Из клеток кишок в их просвет поступает НСОг, а Н+ перемещаются в ток крови.
Н 1 , поступающие в кишки, нейтрализуются НСОз .
28

Рис. 6. Монограмма плазмы крови, пищеварительных соков и пота:
а -нормальная номограмма, |
б—ацидоз, |
в—алкалоз, .' — желудочный сок, |
d — панкреатический |
сок, е—пот |
(Э. Керпель-Фрониус, 1981) |
Поддержание равновесия КОС тесно связано в организме с другими видами обмена и прежде всего с водно-электролитным. Пищеварительный канал, наряду с почками, играет важную роль в поступлении и выделении электролитов. Особенно большая потеря электролитов происходит при рвоте и диарее. Желудочный сок содержит Н* и С1~ больше, чем Na*". В связи с этим рвота является причиной гипохлоремии и гипохлоремического алкалоза. Кишечный сок содержит бикарбонатов и натрия больше, чем хлора, в связи с чем понос является причиной гиперхлоремии и ацидоза.
При ацидозе, в связи с потерей бикарбонатов увеличивается содержание С1~~, что сохраняет электронейтралыюсть. Алкалоз сопровождается накоплением бикарбонатов (из-за усиления реабсорбции последних в почечных канальцах), а количество С1 " уменьшается (рис. 6). Монограмма желудочного сока показывает, что при потере его (рвоте) организм теряет большое количество Η f и С1~, что может привести к состоянию алкалоза. Потеря же панкреатического сока способствует большой потере бикарбонатов и развитию ацидоза.
При рвоте и поносе теряется большое количество калия, что может привести к развитию гипокалиемического алкалоза.
Р о л ь п е ч е н и в п о д д е р ж а н и и КОС . Кровь, оттекающая от кишок, содержит первично недоокисленные продукты типа органических кислот. В печени при достаточном количестве кислорода благодаря циклу Кребса происходит окисление их до конечных продуктов: НгО и СОг (рис. 7). При этом из кишок в печень поступает аммиак (Nbb) и аммония хлорид (NH4CI), из которых синтезируется нейтральное вещество — мочевина.
Благодаря выделительной функции печени с желчью выделяется в кишки большое количество как кислых, так и основных соединений.
Гипоксическое состояние, функциональная неполноценность
29

Рис. 7. Роль печени в поддержании КОС
печеночных клеток (гепатит, цирроз печени) приводят к нарушению окислительных процессов в печени. При этих состояниях в кровоток будут поступать соединения с кислыми свойствами
ибудет развиваться метаболический ацидоз.
Внастоящее время определение показателей кислотно-основ- ного состояния осуществляется по методу Зиггарда — Андерсена на аппарате Аструпа (микрометод Аструпа). В основу метода положено определение истинного рН плазмы и рН этой же среды после сатурации ее газовой смесью, содержащей 4 % и 8 % СОг. Полученные три величины рН являются отправными точками для расчета по номограмме Зиггарда — Андерсена показателей КОС. Основные показатели КОС приведены в табл. 10. Классификация нарушений КОС представлена на рис. 8.
Метаболический ацидоз развивается в результате накопления Н+ в организме или потере НСОз". Первая защита от нагрузки кислотами осуществляется буферной системой бикарбонат / угольная кислота. Происходит связывание Н+ в реакции
На начальных этапах компенсаторные механизмы обеспечивают расщепление угольной кислоты до СОг и НгО, благодаря чему сохраняется нормальным соотношение:
Когда кислотность достигает уровня, который снижает рН крови, в поддержание КОС вступает второй барьер защиты — легочная вентиляция. Развивается компенсаторная легочная гипервентиляция и увеличивается выведение СОг из организма. При снижении рН крови на 0,1 легочная вентиляция увеличивается в 2 раза. При повышении рСО2 на 10 мм рт. ст. вентиляция увеличивается в 4 раза, а при рСОг 70 мм рт. ст.— в 10 раз. Выведение СОг через органы дыхания способствует уменьшению содержания угольной кислоты и поддержанию отношения бикарбонат / угольная кислота на нормальном уровне.
Метаболический ацидоз становится декомпенсированным,
30