

3.5. Механизм действия гормонов
В отличие от ферментов и витаминов гормоны изменяют скорость протекания не какой-то конкретной химической реакции или группы сходных реакций, а затрагивают в обмене веществ некие фундаментальные процессы.
Стероидные гормоны регулируют в клетках-мишенях синтез в первую очередь информационных РНК. Последние немедленно обеспечивают наработку специфических белков, оказывающих влияние на обмен веществ, и, следовательно, на физиологические процессы
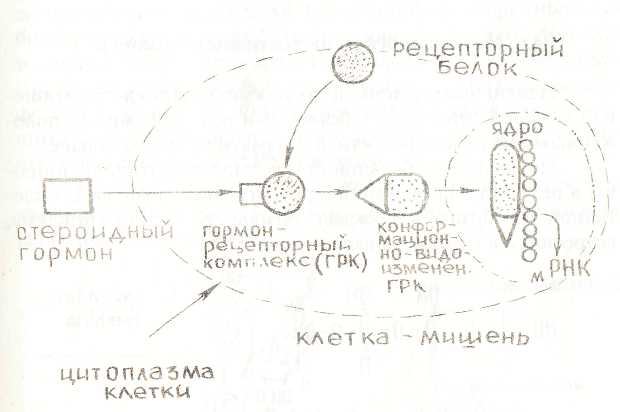
С хроматином* вступает в контакт ГРК, после присоединения стероидного гормона к белку-рецептору гормон часто преобразуется, например, восстанавливается по двойной связи в кольце А, а белок-рецептор изменяет свою конформацию.
В новой конформации ГРК транслоцируется в ядро, где связываете с акцепторным участком хроматина, переводя ДНК в этой зоне хроматина в транскрипционно-активное состояние.
* - Хроматин - суммарный материал хромосом - содержит ДНК, гистоны («-50%) (простые белки), негистоновые белки, небольшое количество РНК
Важно подчеркнуть, что лишь строго определенный стероидный гормон «узнает» свою клетку-мишень, а в ней - свой специфический белок-рецептор. В свою очередь ГРК сугубо избирательно атакует в ядре клетки ту область хроматина, которая содержит фрагмент ДНК, ответственный за синтез индивидуальной мРНК и, следовательно, белка, с только ему присущей функциональной активностью. Белки-рецепторы интенсивно изучаются. Так, рецепторный белок для гидрокортизона имеет М = 67000, а для эстрадиола М = 200000.
3.6. Механизм действия пептидных гормонов
Пептидные гормоны не проникают внутрь клеток-мишеней и взаимодействуют с белковыми рецепторами, расположенными на их поверхности, в плазматической мембране.
Подавляющее большинство гормонов пептидной природы в результате связывания с рецепторным комплексом клеточной мембраны возбуждает активность аденилатциклазы, встроенной в ту же мембрану.
Возникающее при этом соединение - циклический аденозинмонофосфат является тем веществом, которое передает гормональный сигнал метаболическим системам клетки.
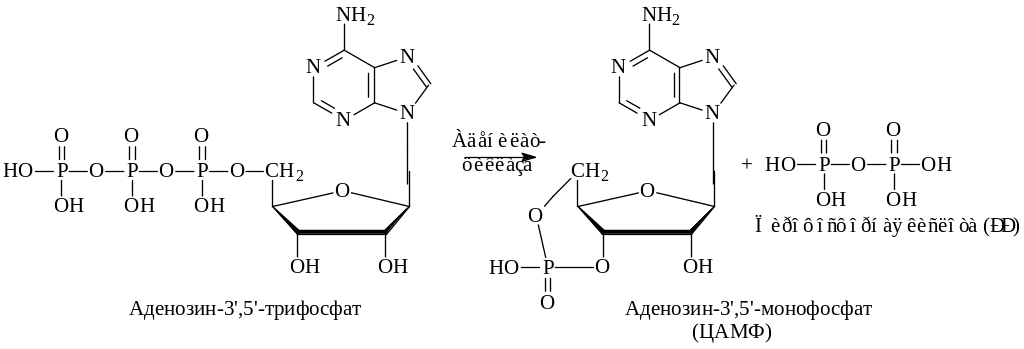
Гормоны (Г1, Г2) присоединяются к специфическим рецепторам мембраны (Р1, Р2).Это присоединение воздействует на мембранный фермент - аденилатциклазу, которая при этом активируется и за счет внутриклеточного АТФ синтезируется цАМФ. цАМФ,в свою очередь, является активатором другого фермента - протеинкиназы. Эта активация осуществляется путем присоединения цАМФ к регуляторной субъединице фермента Р, что приводит к освобождению каталитической субъединицы К. Активированная протеинкиназа катализирует фос-форилирование того или иного белка (фермента, гистона) в зависимости от типа клетки. В результате фосфорилирования биологические свойства белка изменяются. Если это фермент, то он может быть активирован или, наоборот, инактивирован. Другой фермент - фосфопротеинфосфатаза может отщеплять фосфат от белка; цАМФ инактивируется ферментом, находящимся в мембране и в цитозоле - фосфодиэстеразой, которая преобразует цАМФ в АМФ, гидролизуя связь 3'.
Передача гормонального сигнала аденилатциклазе осуществляется в результате работы довольно сложного гормон-рецепторного комплекса.
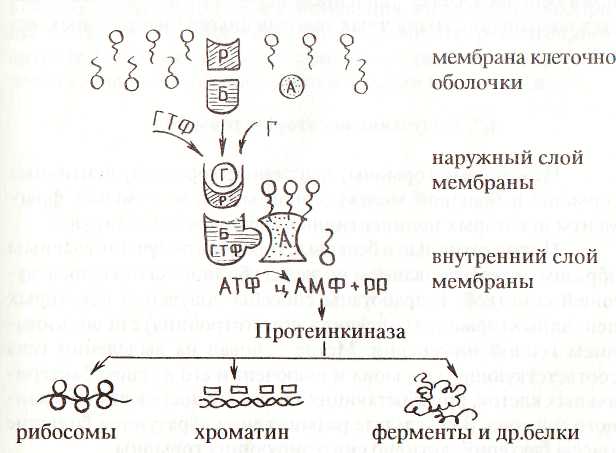
Р - рецептор гормона; Б - регуляторный белок; А - аденилатциклаза; Г - гормон; ГДФ - гуанозиндифосфат; ГТФ - гуанозинтрифосфат
При взаимодействии рецептора с гормоном сигнал передается регуляторному белку, в котором связанная с ним ГДФ заменяется на ГТФ. При этом возбуждается ГТФазная активность, происходит гидролиз ГТФ и выделившаяся энергия используется для кратковременного изменения конформации регуляторного белка и сопряженной с этим аденилатциклазы. Последняя, переходя в активный конформер, ускоряет реакцию биосинтеза цАМФ. После того, как сигнал принят и реализован, аденилатциклазная система приходит в исходное состояние.
Некоторые из гормонов пептидной природы действуют не по аденилатциклазному механизму. Например, инсулин, связываясь с белковым рецептором (М = 300000 гликопротеин, состоит из нескольких субъединиц) плазматической мембраны клетки-мишени, изменяет ее проницаемость. В результате этого усиливается проникновение в клетку субстратов (глюкоза, аминокислоты и др.) и в ней на полную мощность включаются в работу соответствующие ферменты. Аналогичным образом действует окситоцин: образование гормон-рецепторного комплекса сопровождается усилением переноса Са2+, что инициирует сокращение мышечных волокон альвеол и молочных желез.