

Математическое моделирование / Arz_(2010)_Mathematical_&_Computer_Modelling
.pdfТаким образом, анализ применимости уравнений «типа Моно» для двух различных микроорганизмов, растущих на разных субстратах, позволяет сделать следующие выводы:
−уравнения «типа Моно» (уравнения (2.124)−(2.128)) удовлетворительно описывают результаты экспериментов при неизменных концентрациях субстратов и биомассы;
−расчеты динамики накопления биомассы и потребления субстрата, полученные по моделям (2.124)−(2.128) при одних и тех же начальных условиях практически не различимы, что не позволяет сделать вывод о преимуществах какой-либо одной из них.
Возможность использования моделей при различных начальных концентрациях субстрата и биомассы. В преды-
дущем разделе были проанализированы возможности использования моделей «типа Моно» для описания кинетики биосинтеза при одинаковых концентрациях субстрата и биомассы. Для большей общности было бы желательно провести подобный анализ и для широких диапазонов изменения концентраций. Для описания кинетики мы использовали следующую модель:
dX =μX − KD X |
(2.129) |
||||
dt |
|
|
|
|
|
μ = μ (S ) |
|
|
(2.130) |
||
dS = |
− |
1 |
μ |
, |
(2.131) |
|
|||||
dt |
|
Y |
X |
||
|
|
|
|
||
где слагаемое KDX отвечает за отмирание микроорганизмов (KD – константа), а удельная скорость роста (2.130) зависит от концентраций субстрата по формуле Моно (2.125). Вначале были определены значения кинетических констант модели для каждого из шести опытов в отдельности. Результаты вычислений приведены в табл. 2.13.
180
|
|
|
|
|
|
|
|
Таблица 2.13 |
|
|
|
|
Кинетические константы для модели Моно |
|
|||||
|
|
|
|
|
|
|
|
|
|
|
Опыт |
Начальные |
|
Константы модели |
|
Значение Ф |
|||
|
X0 |
|
S0 |
μm, ч−1 |
Y, г/г |
||||
|
|
KS, г/л |
KD, ч−1 |
||||||
|
|
Концентрации, г/л |
|
|
|
|
|||
|
|
|
|
|
|
|
|
|
|
|
1 |
11,7 |
|
1,96 |
0,135 |
1,4966 |
0,027 |
1,3128 |
0,29998 |
|
2 |
6,28 |
|
1,96 |
0,2326 |
1,784 |
0,0394 |
1,2914 |
0,26929 |
|
3 |
2,7 |
|
1,96 |
0,6028 |
2,5998 |
0,10777 |
2,0418 |
0,12533 |
|
4 |
9,4 |
|
1,98 |
0,1058 |
1,1872 |
0,0242 |
1,3264 |
0,25974 |
|
5 |
7,2 |
|
1,98 |
0,15541 |
0,76919 |
0,05929 |
2,35995 |
0,05900 |
|
6 |
2,0 |
|
0,55 |
0,47742 |
0,79824 |
0,04926 |
1,45474 |
0,00393 |
Выполненные расчеты показывает, что модель (2.129)−(2.131) удовлетворительно описывает каждый из шести опытов в отдельности (рис. 2.91). Максимальные приведенные погрешности для биомассы и субстрата составляют около 1 и 3 %. Вместе с тем значения всех без исключения кинетических констант существенно различаются в разных опытах. Это ставит под сомнение возможность получения единой модели, адекватной рассматриваемому объекту в широких диапазонах концентраций.
По этой причине была предпринята попытка получить значения кинетических констант, справедливых для всех шести опытов (табл. 2.13) одновременно. Были получены следующие значения: μm = 0,07088 ч−1, KS=0,09944 г/л, KD=0,02053 ч–1,
Y = 1,02193 г/г. Значение критерия Ф для всех шести опытов в этом случае составляет 7,64872. Из рис. 2.92 следует, что модель (2.129)−(2.131) с указанными значениями констант не позволяет получить удовлетворительного описания экспериментальных данных. Наиболее существенные погрешности наблюдаются в описании концентраций субстрата в опытах 1, 3−6 (рис. 2.92 а, c−f) и концентраций биомассы в опытах 2 и 3 (рис. 2.92 b, c).
Таким образом, из анализа расчетных и экспериментальных данных, приведенных в этом разделе можно сделать следующие выводы:
− модель «типа Моно» удовлетворительно описывает кинетику роста микроорганизмов и потребления субстрата при конкретных начальных концентрациях;
181
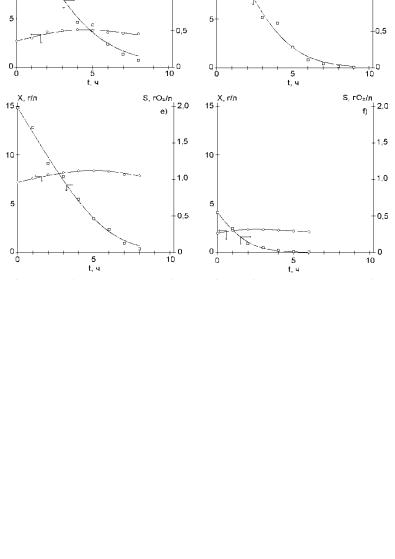
Рис. 2.91. Описание экспериментальных данных для кормовых дрожжей на комплексном субстрате моделью (2.129)−(2.131): a) X0 = 11,7; S0 = 1,96; b) X0 = 6,28; S0 = 1,96; c) X0 = 2,7; S0 = 1,96; d) X0 = 9,4; S0 = 1,98; e) X0 = 7,2;
S0 = 1,98; f) X0 = 2,0; S0 = 0,55. Точки – экспериментальные данные, линия – расчет по соответствующей модели
182

Рис. 2.92. Описание экспериментальных данных для кормовых дрожжей на комплексном субстрате моделью (2.129) − (2.131): a) X0 = 11,7; S0 = 1,96; b) X0 = 6,28; S0 = 1,96; c) X0 = 2,7; S0 = 1,96; d) X0 = 9,4; S0 = 1,98; e) X0 = 7,2; S0 =
1,98; f) X0 = 2,0; S0 = 0,55. Точки – экспериментальные данные, линия – расчет по соответствующей модели
183
− эта же модель не позволяет проводить адекватное описание экспериментальных данных при различных начальных концентрациях биомассы и субстрата, изменяющихся в широких диапазонах.
Моделирование режимов работы биохимических реакторов при использовании кинетики «типа моно». Математи-
ческая модель биохимического реактора непрерывного типа с идеальным смешением по жидкой фазе в случае с одним лимитирующим субстратом может быть записана в виде:
dX |
= μ (S) X − DX |
|
(2.132) |
||||
dt |
|
||||||
|
|
μ (S)X |
|
|
|||
dS |
|
= D (S0 |
− S) − |
, |
(2.133) |
||
dt |
Y |
||||||
|
|
|
|
||||
где Х – концентрация биомассы в реакторе; S, S0 – концентрация субстрата в реакторе и начальная концентрация субстрата на входе в реактор; Y – экономический коэффициент; μ – удельная скорость роста биомассы; D = F/V – удельное разбавление; F – расход субстрата через биохимический реактор; V – объем биохимического реактора. Соответственно для кинетических зависимостей μ(S), представленных в табл. 2.12 уравнениями (2.122)−(2.128), модель проточного биохимического реактора (2.132)−(2.133) может быть записана в одной из следующих форм.
1. μ(S)=k: |
|
|
|
|
|
||
|
dX |
= kX − DX |
(2.134) |
||||
|
dt |
|
|
|
|
|
|
|
dS |
= D (S0 |
− S) − |
kX |
(2.135) |
||
|
dt |
Y |
|||||
|
|
|
|
|
|
||
2. μ(S)=kS: |
|
|
|
|
|
||
|
dX |
= kSX − DX |
(2.136) |
||||
|
dt |
|
|
|
|
|
|
|
dS |
= D (S0 |
− S) − |
kSX |
|
(2.137) |
|
|
dt |
|
|
||||
|
|
|
|
Y |
|
|
|
184
3. μ(S)=kSn: |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|||||||
|
dX |
|
= kS n X − DX |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|||||||||||
|
dt |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||
|
dS |
|
= D (S0 − S) − |
kS n X |
|
|
|
|
|
|
|
|
|
|
|||||||||||||||||
|
dt |
|
|
|
|
Y |
|
|
|
|
|
|
|
|
|
|
|
||||||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|||||
4. μ |
(S)= μ |
|
|
|
|
S |
|
|
: |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||||||||
|
|
|
|
|
|
|
m S + KS |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|||||||
|
dX |
|
= μ |
|
|
|
|
|
S |
X − DX |
|
|
|
|
|
|
|
|
|
|
|
||||||||||
|
dt |
|
m S |
|
+ KS |
|
|
|
|
|
|
|
|
|
|
|
|||||||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||||||||
|
dS |
= D (S |
|
|
− S) −μ |
|
|
|
|
|
S |
|
|
|
X |
|
|
|
|
||||||||||||
|
dt |
|
|
|
|
|
|
|
|
|
|
Y |
|
|
|
|
|||||||||||||||
|
|
|
|
|
|
0 |
|
|
|
|
|
m S + KS |
|
|
|
|
|||||||||||||||
5. μ (S )= μm |
Sn |
|
: |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|||||||||||||
S n + KS |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|||||||||||||||
|
dX |
|
= μ |
|
|
|
|
|
Sn |
|
X − DX |
|
|
|
|
|
|
|
|
|
|
||||||||||
|
dt |
|
m Sn |
|
|
|
|
|
|
|
|
|
|
|
|
|
|||||||||||||||
|
|
|
|
+ KS |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||||||||||
|
dS |
|
= D (S |
|
|
− S) −μ |
|
|
|
|
|
S n |
|
|
|
|
|
|
X |
|
|
|
|||||||||
|
dt |
0 |
|
m S n |
+ KS |
|
|
|
Y |
|
|
|
|||||||||||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||||||||||||
6. μ (S)= μm (1−e−S / k ): |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|||||||||||||||||
|
dX |
|
= μm (1−e−S / k )X − DX |
|
|
|
|
|
|
|
|
|
|
|
|||||||||||||||||
|
dt |
|
|
|
|
|
|
|
|
|
|
|
|
||||||||||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||
|
dS |
= D (S0 − S) −μm (1−e−S / k ) |
|
|
X |
|
|
|
|
||||||||||||||||||||||
|
dt |
|
|
Y |
|
|
|
|
|||||||||||||||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||||
7. μ (S)=μm |
|
|
|
S |
|
|
|
|
|
: |
|
|
|
|
|
|
|
|
|
|
|||||||||||
S + KS +kS2 |
|
|
|
|
|
|
|
|
|
|
|||||||||||||||||||||
|
dX |
|
= μ |
|
|
|
|
|
|
S |
|
|
|
|
|
|
|
X − DX |
|
|
|
|
|
|
|
|
|||||
|
dt |
|
m S |
|
+ KS + kS2 |
|
|
|
|
|
|
|
|
|
|||||||||||||||||
|
|
|
|
|
|
|
|
S |
|
|
|
|
|
|
|
X |
|||||||||||||||
|
dS |
= D (S |
|
|
− S) −μ |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||||||||||||
|
dt |
0 |
m S + KS + kS 2 |
|
Y |
||||||||||||||||||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|||||||||||||||||||
(2.138)
(2.139)
(2.140)
(2.141)
(2.142)
(2.143)
(2.144)
(2.145)
(2.146)
(2.147)
185

Учитывая, что при использовании зависимости μ(S)=k, не удается получить высокого качества описания экспериментальных данных (см. табл. 2.12), мы не рассматривали в дальнейших рассуждениях модель биореактора, построгенную на основе уравнений (2.134) и (2.135).
Уравнения для статических режимов могут быть получены, если положить в уравнениях (2.136)−(2.147) dX/dt=dS/dt=0. Тогда получим следующие зависимости для расчета концентраций X и S в статическом режиме на выходе из реактора (внутри реактора).
1. μ(S)=kS:
S = Dk
X= S0 − D Y
k
2.μ(S)=kSn:
1
S = D nk
|
|
|
1 |
|
|
|
X = S |
|
D n |
|
Y |
||
0 |
− |
|
|
|||
|
||||||
|
|
k |
|
|
||
|
|
|
|
|
|
|
3. μ (S )= μm S +SKS :
S = KS D
μm − D
X = |
S |
0 |
− |
KS D |
Y |
|
|||||
|
|
|
|
|
|
|
|
|
|
μm − D |
|
(2.148)
(2.149)
(2.150)
(2.151)
(2.152)
(2.153)
186

4. μ (S)= μm |
|
|
|
Sn |
: |
||||||
Sn + KS |
|
||||||||||
|
|
|
|
|
1 |
|
|
|
|
||
|
|
KS D |
|
|
n |
|
|
|
|||
S = |
|
|
|
|
|
||||||
|
|
|
|
|
|||||||
|
|
|
|
|
|
||||||
|
μm − D |
|
|||||||||
|
|
|
|
|
|
|
n |
||||
|
|
|
|
|
1 |
|
|||||
X = |
|
|
|
KS D |
|
|
|||||
|
|
||||||||||
|
S0 − |
|
|
|
|
|
Y |
||||
|
μm − D |
|
|||||||||
|
|
|
|
|
|
|
|
|
|
|
|
5. μ (S )= μm (1 − e−S / k ):
S = −k ln μm − D μm
X = S0 +k ln μmμ− D Y
m
6. μ (S)= μm S + KS +kS2 :
S
S = (μm − D)± (μm − D)2 −4D2k KS |
||||||
|
|
|
2D k |
|
|
|
|
|
|
2 |
− |
2 |
|
X = |
|
− |
(μm − D)± (μm − D) |
4D |
k KS |
|
S0 |
2D k |
|
|
Y |
||
|
|
|
|
|
|
|
(2.154)
(2.155)
(2.156)
(2.157)
(2.158)
(2.159)
Заметим, что при использовании кинетического уравнения (2.128) табл. 2.12 при расчете X и S по уравнениям (2.158)−(2.159) имеют место два статических состояния. Анализ условий функционирования проточного биореактора позволяет сделать вывод о том, что интересующий нас режим наблюдается при использовании в уравнениях (2.158)−(2.159) знака «–».
Для анализа режимов работы биохимических реакторов наибольшее значение имеет изучение зависимости X(D), как основного технологического параметра, а также удельной продуктивности биореактора по биомассе Q(D) = D. X (D).
187

Рис. 2.93. Зависимости концентрации биомассы X от величины удельного разбавления D в статическом режиме при использовании различных моделей кинетики. Буквы соответствуют уравнениям: a) − (2.149); b) − (2.151); c) − (2.153); d) − (2.155); e) − (2.157); f) − (2.159). Начальные концентрации субстрата со-
ставляют: I − 0,5 г/л; II − 1 г/л; III − 1,5 г/л; IV − 2 г/л; V − 2,5 г/л; VI − 3 г/л
188
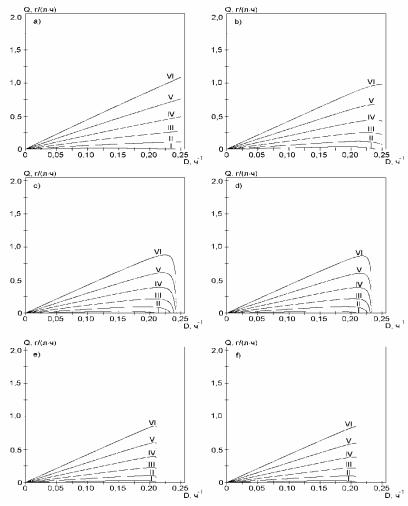
Рис. 2.94. Зависимости удельной продуктивности Q от величины удельного разбавления D в статическом режиме при использовании различных моделей кинетики. Буквы соответствуют уравнениям: a) − (2.149); b) − (2.151); c) − (2.153); d) − (2.155); e) − (2.157); f) − (2.159). Начальные концентрации субстрата составляют: I − 0,5 г/л; II − 1 г/л; III − 1,5 г/л; IV − 2 г/л; V − 2,5 г/л; VI − 3 г/л.
189