

Математическое моделирование / Arz_(2010)_Mathematical_&_Computer_Modelling
.pdfНа рис. 2.87 слева изображен внешний вид модельной популяции (двойного кластера) со следующими значениями параметров и начальных условий: начальное количество питательного вещества – 200; скорость потребления питательного вещества – 1; скорость восполнения питательного вещества – 0; коэффициент диффузии – 0,05; среднее значение максимальной продолжительности жизни – 100; среднее значение времени между делениями микроорганизма – 10. Начальное положение клеток – 2 ячейки с координатами (24,67) и (70,38). Начальный возраст клеток выбирали исходя из равномерного распределения от 0 до среднего значения максимальной продолжительности жизни.
На этом же рисунке справа изображен внешний вид популяции плесневых грибов Aspergillus полученной при помощи пассивной воздушной аэрации в помещении.
Рис. 2.87. Слева – внешний вид популяции, полученный по модели [10] при Q0
= 200; Q- = 1; Q+ = 0; η = 0,05; τl = 100; τd = 10 в различные моменты времени: a) – 2, b) – 42, c) – 97, d) – 135, e) – 169, f) – 190, g) – 211, h) – 232, i) – 254, j) – 275, k) – 296, l) – 317, m) – 334, n) – 355 итераций; одна итерация равняется
852 с. Справа – формы колоний плесневых грибов Aspergillus полученные в эксперименте в различные моменты времени: a) – 0,5, b) – 10, c) – 23, d) – 32, e) – 40, f) – 45, g) – 50, h) – 55, i) – 60, j) – 65, k) – 70, l) – 75, m) – 79, n) – 84 ч
после засева; размер области 3 × 3 см
170

Сравнивая изображения, полученные по модели и в эксперименте на рис. 2.86 и 2.87 можно сделать вывод об их морфологическом соответствии.
Соответствие наблюдается как в размерах, так и в форме популяции, небольшие отклонения от округлых форм в модельной популяции объясняются дискретностью сетки и учетом взаимодействия только с четырьмя ближайшими ячейками. Столкновения областей распространения рис. 2.87 имеют много общего для экспериментальной и модельной популяции, и имеют одинаковую динамику. Внутри областей распространения в эксперименте имеются темные пятна, связанные с малоактивными областями, также эти области хорошо заметны и в модели и развиваются похожим образом.
Таким образом, можно утверждать, что математическая модель роста биологических объектов на плоскости, разработанная нами ранее позволяет адекватно описывать кинетику и морфологию таких объектов.
2.6. Математические модели кинетики микробиологического синтеза: возможности использования и новые подходы к разработке1
Существующие подходы к моделированию кинетики микробиологического синтеза. В настоящее время для описа-
ния кинетики микробиологического синтеза, под которой обычно понимают динамику потребления субстрата, продукции биомассы и метаболитов используется большое количество математических моделей. Большинство из них, такие как модели Моно, Халдейна, Тессье и др. имеют вполне определенный физический смысл. Так, например, модель Моно опирается на допущение о существовании в системе биохимической реакции с промежуточным фермент-субстратным комплексом вида:
1 Арзамасцев А. А., Андреев А. А. О возможности использования различных моделей кинетики биосинтеза // Биофизика. – 2001. – Т. 46. – Вып. 6. –
С. 1048-1061.
171
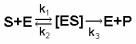
(2.117)
Это обстоятельство делает ее по форме схожей с хорошо известной в биофизике моделью Микаэлиса–Ментен.
Существует достаточно большое число математических моделей, позволяющих описывать кинетику роста биомассы и ассимиляции субстрата. Все они обычно выражаются зависимостями:
dX |
= μX |
|
|
|
|
(2.118) |
||
dt |
|
|
|
|
||||
|
(S) |
|
|
|
|
|||
μ = μ |
|
|
|
(2.119) |
||||
dS |
= |
− |
1 |
|
dX |
, |
(2.120) |
|
dt |
Y |
dt |
||||||
|
|
|
|
|
||||
где Х − концентрация биомассы; S − концентрация субстрата; Y − экономический коэффициент, показывающий выход биомассы с единицы субстрата; μ − удельная скорость роста биомассы. Для получения решения система (2.118)−(2.120) должна быть дополнена соответствующими начальными условиями. В дальнейшем модели, которые обобщенно могут быть представлены в виде уравнений (2.118)−(2.120) мы и будем называть моделями «типа Моно». Точность аппроксимации экспериментальных данных уравнениями (2.118)−(2.120) в основном зависит от правильного выбора зависимости удельной скорости роста биомассы от концентрации субстрата (2.119). К уравнениям такого типа могут быть отнесены зависимости Моно, Мозера, Тессье, Моно-Иерусалимского и ряд других. На рис. 2.88 показан общий вид зависимостей μ(S), характерный для различных моделей «типа Моно». Из этого рисунка видно, что при малых концентрациях субстрата (0 ≤ S ≤ S1) может быть в равной степени использована любая из рассмотренных моделей (см. табл. 2.12). Лимитирование удельной скорости роста микроорганизмов субстратом учитывают лишь модели Моно, Мозера, Тесье и Халдейна (0 ≤ S ≤ S2). Ингибирование удельной скорости роста биомассы большими концентрациями субстрата из рассматриваемых моделей описывает лишь модель Халдейна.
172
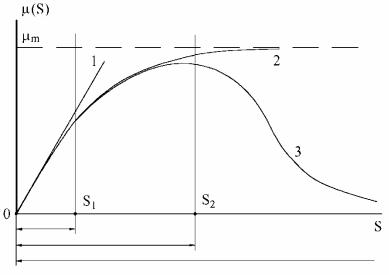
Рис. 2.88. Области применения различных кинетических моделей: 1 – модель вида μ (S)=kS, рабочий диапазон 0 ≤ S ≤ S1; 2 – модель вида μ (S)=μmS/(S+KS) или μ (S)=μmSn/(Sn+KS), рабочий диапазон 0 ≤ S ≤ S2; 3 – модель вида μ (S)=μmS/(S+KS+kS2) применяется в широком диапазоне концентраций S
Модели «типа Моно» имеют ряд достоинств: относительно простой вид самих уравнений, небольшое количество коэффициентов и возможность их идентификации по экспериментальным данным, хорошо отработанные процедуры получения решений.
Однако, эти модели не учитывают целый ряд явлений, сопутствующих биосинтезу: диффузионные процессы транспорта субстратов в зону реакции и отвода продуктов, обусловленные гидродинамическими режимами в биореакторе; возрастную, фазовую, физико-химическую, морфологическую и другие виды гетерогенности самой популяции микроорганизмов.
По этой причине вопрос о применимости моделей «типа Моно» остается открытым. С одной стороны данные модели все-таки используются многими авторами в ограниченных диапазонах описываемых переменных; с другой – имеется достаточно выраженная критика в их адрес.
173
Одновременно с широким использованием моделей «типа Моно» многие авторы применяют и другие подходы к описанию кинетики микробиологического синтеза.
Так, в соответствии с уравнением Ферхюльста рост популяции микроорганизмов в ограниченном пространстве характеризуется «S-образной» кривой с насыщением. По мнению некоторых авторов, эта модель учитывает внутривидовую конкуренцию. Однако анализ этого уравнения, проведенный Ч. Файси, показал, что оно является лишь частным случаем модели Моно при низких концентрациях субстрата.
Во многих случаях имеются попытки учесть различные виды гетерогенности микробных популяций при их росте.
Таким образом, результаты многочисленных исследований говорят о довольно разнообразных подходах к построению моделей роста микробиологических популяций. Вместе с тем, в настоящее время так и не выработано единой точки зрения относительно того базиса, на который должен опираться процесс построения такой модели. Не вполне ясно, рассмотрение каких явлений: биохимических (метаболитические пути, механизм реакции, процессы регуляции), физических (диффузионные процессы) или биологических (возрастная, морфологическая и др. виды гетерогенности популяции) наиболее существенно для ее построения.
По этой причине вопрос о разработке модели макрокинетики для процессов микробиологического синтеза, ее базисе и структуре является актуальным. В этой связи представляется важным провести тщательный анализ возможностей использования уже существующих на данный момент, особенно часто используемых моделей «типа Моно», а также и формирование базисов для построения новых моделей.
В рамках рассматриваемой цели необходимо дать ответы на следующие вопросы:
−возможно ли использование моделей «типа Моно» для различных субстратов и микроорганизмов при одинаковых или близких их концентрациях;
−как выбрать лучшую модель;
174
−можно ли использовать эти модели при различных начальных концентрациях субстрата и биомассы;
−можно ли на основе моделей «типа Моно» рассчитывать основные характеристики биореакторов; насколько велики погрешности таких расчетов;
−как и с каких позиций должна разрабатываться математическая модель кинетики микробиологического синтеза.
Возможность использования моделей «типа Моно» в узких диапазонах концентраций. Для оценки применимости мо-
делей использовались экспериментальные данные по кинетике биосинтеза на различных субстратах и для разных микроорганизмов.
Для идентификации параметров математических моделей и их сравнения разработана специальная программа на языке Turbo Pascal 7.0. Программа реализует безградиентный метод покоординатного спуска и позволяет рассчитывать по экспериментальным данным коэффициенты моделей, минимизирующие взвешенную сумму квадратов отклонений экспериментальных и расчетных значений:
Ф = ∑m |
β1 (X (ti )− X iэ )2 |
+β2 (S(ti )− Siэ )2 → min |
(2.121) |
i=1 |
|
|
|
где β1, β2 − весовые коэффициенты; Siэ, Xiэ − эксперименталь-
ные значения концентраций субстрата и микроорганизмов; S (ti), X (ti) − расчетные значения этих концентраций.
Объект 1. Экспериментальные данные по росту биомассы бактерий рода Pseudomonas на комплексном субстрате при 30 0С показаны на рис. 2.89. На их основе проводилось сравнение различных математических моделей вида (2.118)−(2.120), зависимости μ(S) для которых приведены в табл. 2.12. Параметры каждой из моделей выбирались исходя из минимального значения критерия (2.121). Значения этого критерия и кинетических констант для различных случаев приведены в табл. 2.12.
175
Анализ рис. 2.89 позволяет сделать следующие выводы. Все математические модели, за исключением рис. 2.89 а (уравнение (2.122), табл. 2.12) удовлетворительно описывают экспериментальные данные. Максимальные приведенные погрешности для биомассы и субстрата составляют 5,2 и 2,8 %, что соответствует сумме квадратов отклонений, подсчитанных по уравнению (2.121) в 1,46. Визуальная оценка результатов расчетов по моделям не позволяет выявить преимущества какой-либо из них. В соответствие с табл. 2.12, сумма квадратов отклонений расчетных и экспериментальных значений для объекта 1 в случае использования уравнений (2.124)−(2.128) также различается незначительно. Простые вычисления показывают, что различия у использованных моделей (2.124)−(2.128) соответствуют приведенной погрешности примерно в 0,12 %, что существенно меньше погрешности экспериментальных данных. Поэтому невозможно выбрать лучшую модель из уравнений (2.124)−(2.128) табл. 2.12, используя в качестве критерия уравнение (2.121). Однако, на основе критерия (2.121) могут быть отброшены две наиболее неподходящие модели (уравнения (2.122), (2.123) табл. 2.12.
Объект 2. Аналогично объекту 1 проводилось сравнение различных математических моделей, уравнения которых приведены в табл. 2.12, на основе экспериментальных данных по кинетике роста микроорганизмов Paecilomyces Variotii на отходах сульфата-целлюлозы. Результаты приведены на рис. 2.90 и в табл. 2.12.
Анализ рис. 2.90 позволяет сделать следующие выводы. Все математические модели, за исключением рис. 2.90 a, b (уравнения (2.122), (2.123) табл. 2.12) удовлетворительно описывают экспериментальные данные. Максимальные приведенные погрешности для биомассы и субстрата составляют 6,1 и 1,9 %, что соответствует сумме квадратов отклонений, подсчитанных по уравнению (2.121) в 0,00416. Визуальная оценка результатов расчетов по моделям так же, как и для объекта 1, не позволяет выявить преимущества какой-либо из них. В соответствие с табл. 2.12, сумма квадратов отклонений расчетных и экспериментальных значений для объекта 2 в случае использования уравнений (2.124)−(2.128) также различается незначительно.
176
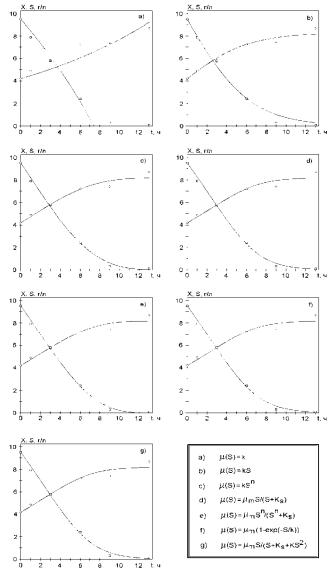
Рис. 2.89. Описание экспериментов для Pseudomonas на комплексном субстрате различными кинетическими моделями. Концентрация субстрата выражена в гO2/л (по БПК5). Точки – экспериментальные данные, линия – расчет по соответствующей модели
177
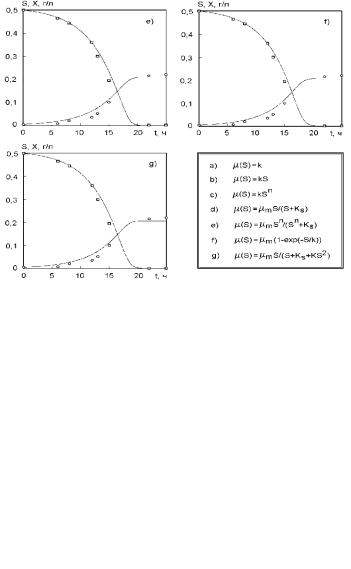
Рис. 2.90. Описание экспериментов для Paecilomyces Variotii на отходах суль- фата-целлюлозы различными кинетическими моделями. Концентрация субстрата выражена в гO2/л (по БПК5). Точки – экспериментальные данные, линия – расчет по соответствующей модели
178
Вычисления показывают, что различия у использованных моделей (2.124)−(2.128) соответствуют приведенной погрешности примерно в 0,3 %, что также существенно меньше погрешности экспериментальных данных. Поэтому невозможно выбрать лучшую модель из уравнений (2.124) − (2.128) табл. 2.12 используя в качестве критерия уравнение (2.121). Наиболее неподходящие модели (уравнения (2.122), (2.123) табл. 2.12) так же, как и для объекта 1 могут быть отброшены на основе критерия (2.121). Значение n в модели (2.126) мало отличается от 1 для обоих объектов, что служит указанием на существование в системе механизма, работающего в соответствие со схемой (2.117).
|
|
|
|
|
|
|
|
|
|
|
|
Таблица 2.12 |
||
|
Сравнение различных кинетических моделей |
|
||||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Уравнение |
Значения коэффициен- |
|
|
||
|
Кинетическая |
тов, доставляющих |
Значение |
|||||||||||
|
|
минимальное значение |
критерия Ф |
|||||||||||
|
модель |
|
||||||||||||
|
|
критерию Ф* |
|
|
||||||||||
|
|
|
|
|
|
|
|
|
|
Объект 1 |
Объект 2 |
Объект 1 |
Объект 2 |
|
μ(S)=k |
|
|
|
|
|
|
(2.122) |
k=0,0606 |
k=0,16 |
2,87729 |
0,00864 |
|||
|
|
|
|
|
|
Y=0,2508 |
Y=0,175 |
|||||||
|
|
|
|
|
|
|
|
|
|
|
|
|||
μ(S)=kS |
|
|
|
|
|
|
(2.123) |
k=0,01647 |
k=0,505 |
1,45950 |
0,01246 |
|||
|
|
|
|
|
|
Y=0,4327 |
Y=0,435 |
|||||||
|
|
|
|
|
|
|
|
|
|
|
|
|||
μ(S)=kSn |
|
|
|
|
|
|
|
k=0,02663 |
k=0,225 |
|
|
|||
|
|
|
|
|
|
(2.124) |
n=0,70018 |
n=0,09 |
0,82140 |
0,00394 |
||||
|
|
|
|
|
|
|
|
|
|
Y=0,41816 |
Y=0,405 |
|
|
|
μ |
(S )= μm |
|
|
S |
|
μm=0,2048 |
μm=0,245 |
|
|
|||||
|
|
|
|
|
|
(2.125) |
KS=6,8726 |
KS=0,075 |
0,86708 |
0,00416 |
||||
S |
+ KS |
|||||||||||||
|
|
|
|
Y=0,4196 |
Y=0,405 |
|
|
|||||||
|
(S)= μm |
|
|
Sn |
|
|
μm=0,509 |
μm=0,235 |
|
|
||||
μ |
|
|
(2.126) |
KS=16,2162 |
KS=0,055 |
0,82192 |
0,00406 |
|||||||
S |
n |
+ KS |
n=0,7212 |
n=0,985 |
||||||||||
|
|
|
|
|
Y=0,4172 |
Y=0,405 |
|
|
||||||
μ (S )= μm (1 − e−S / k ) |
(2.127) |
μm=0,138 |
μm=0,21 |
0,88839 |
0,00367 |
|||||||||
k=4,997 |
k=0,07 |
|||||||||||||
|
|
|
|
|
|
|
|
|
|
Y=0,4202 |
Y=0,405 |
|
|
|
μ (S )= μm |
|
|
|
S |
|
μm=0,2104 |
μm=0,272 |
|
|
|||||
|
|
|
(2.128) |
KS=7,17536 |
KS=0,072 |
0,86833 |
0,00410 |
|||||||
S + KS + kS 2 |
|
k=0,00073 |
k=0,323 |
|||||||||||
|
|
|
|
|
|
|
|
|
|
Y=0,41997 |
Y=0,403 |
|
|
|
* − размерность кинетических констант соответствует измерению концентраций в г/л и удельной скорости роста в ч−1
179