

Математическое моделирование / Arz_(2010)_Mathematical_&_Computer_Modelling
.pdfнейный рост популяции микроорганизмов во время аутостабилизации температуры, образование температурных колебаний внутри биохимического реактора в процессе саморегулирования температуры. Математическая модель описывала процесс, как при наличии саморегулирования температуры, так и без него.
Целью данного раздела является изучение процесса саморегулирования температуры в популяции микроорганизмов в непрерывном режиме. Математическая модель, которая использована в данной работе, была описана в разделе 2.2.
Исследовали поведение объекта в условиях различных температур во входном потоке, различных внешних температур и различных концентрациях субстрата во входном потоке.
На рис. 2.41 представлена зависимость температуры в биореакторе от времени при различных значениях температуры во входном потоке. Из рисунка видно, что аутостабилизация температуры в непрерывном режиме имеет место не во всех случаях. В случае 1 (рис. 2.41), начиная с нулевой температуры во входном потоке, что фактически соответствует замерзанию жидкости, саморегулирование уже наблюдается. Однако выход реактора на режим аутостабилизации достаточно длителен и составляет около 10 часов. Время выхода на режим аутостабилизации сокращается по мере увеличения температуры во входном потоке. Также видно, что в некоторых случаях, в частности при температурах во входном потоке 0–20 ºС, наблюдается первоначальное снижение температуры в реакторе, и только после того как биологический объект выходит на активный режим по скорости процесса, температура вновь поднимается. В то же время при высоких температурах, в частности 30 ºС и выше, температура начинает повышаться сразу. Наблюдается перерегулирование, т. е. сначала температура превышает оптимальную температуру в биореакторе примерно на 1,5–2 ºС, а затем она выходит на заданный уровень. При температурах 50 ºС и выше биологический объект «не справляется» с задачей саморегулирования, и наблюдаемая в реакторе температура превышает температуру саморегулирования, составляющую в данном случае 40,5 ºС. Однако на кривых 5, 6 и 7 также заметна тенденция биологического объекта к выравниванию температуры. Таким
110
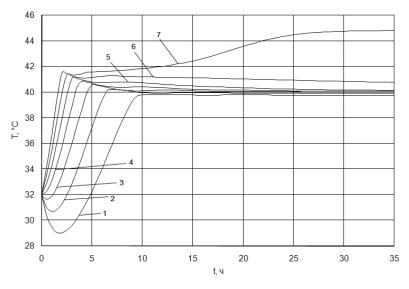
Рис. 2.41. Зависимость температуры в биореакторе от времени при различных значениях температуры во входном потоке. При Text = 33 ºC; D = 0.07 1/ч; Sin = 50 г/л. Номера линий соответствуют различным значениям Tin: 1–0, 2–10, 3–20, 4–30, 5–40, 6–50, 7–60 ºC.
образом можно сделать заключение, что при довольно значительных изменениях температуры во входном потоке, от 0–50 ºС, биологический объект обладает способностью к самопроизвольному выравниванию этого фактора.
На рис. 2.42 показано как меняется температура в реакторе при различных температурах вне реактора. Видно, что при значении внешней температуры 5 ºС биологический объект не справляется с ее выравниванием, хотя такое стремление есть в промежутке 5–15 часов, но затем биологический объект остывает. На кривой 2 рис. 2.42 видно, что вначале температура биологического объекта так же начинает снижаться. Однако по мере выхода ферментных систем организма на заданный режим биологический объект справляется с неблагоприятными воздействиями внешней среды и выравнивает температуру до заданного уровня. Такая же тенденция наблюдается и при температурах
111

20–30 ºС При больших температурах биологический объект стремится выравнивать температуру в сторону снижения. Это заметно на кривых 4–6 при температурах внешней среды 30–
50 |
ºС. На кривой 7, когда температура внешней среды более |
60 |
ºС, видно, что биологический объект не справляется с задан- |
ной задачей и температура в реакторе постепенно начинает повышаться.
На рис. 2.43 показана зависимость температуры в биореакторе от времени при различных значениях концентрации субстрата во входном потоке. Видно, что объект имеет способность к саморегулированию при концентрации субстрата, начиная с 20 г/л, и кривые имеют абсолютно идентичный характер. Одна-
ко, при низких значениях концентрации субстрата |
(10 г/л), |
в результате лимитирования скорости процесса и |
скорости |
Рис. 2.42. Зависимость температуры в биореакторе от времени при различных значениях внешней температуры. При Tin = 20 ºC; D = 0.07 1/ч; Sin = 50 г/л. Номера линий соответствуют различным значениям Text: 1–5, 2–10, 3–20, 4–30, 5–40, 6–50, 7–60 ºC
112

тепловыделения, биологический объект сначала выходит на заданный температурный режим, а затем ввиду ограниченности скорости роста и тепловыделения, температура в реакторе начинает снижаться и достигает уровня не соответствующего температуре аутостабилизации. В других случаях при концентрации субстрата 20–50 г/л во входном потоке объект выходит на статический режим и поддерживает температуру.
Таким образом, в данном разделе осуществлен анализ и исследование математической модели для непрерывного режима. Показано, что режим саморегулирования температуры проявляется здесь намного ярче, чем в периодическом процессе. По крайней мере, температура устанавливается и может сохраняться неопределенно долго, поскольку непрерывный режим не связан с полным исчерпанием субстрата.
Рис. 2.43. Зависимость температуры в биореакторе от времени при различных значениях концентрации субстрата во входном потоке. При Text = 33 ºC; Tin = 20 ºC; D = 0.07 1/ч. Номера линий соответствуют различным значениям
Sin: 1–10, 2–20, 3–30, 4–40 г/л
113

2.4. Морфологические модели развития биологических популяций1
Целью данного раздела является построение дискретной модели роста микроорганизмов на плоскости, позволяющей наблюдать морфологические изменения.
Выбор в качестве модели дискретного представления объекта обусловлен следующими причинами:
–вид реальной колонии микроорганизмов, растущей на плоскости, представляет собой дискретную картину, состоящую из отдельных элементов – клеток; поэтому дискретное представление в большей степени адекватно реальному объекту, чем непрерывное;
–при таком представлении возможно рассмотрение процессов для каждой клетки, в то время как в непрерывной модели можно говорить лишь об осредненных характеристиках колонии или ее части;
–дискретное представление объекта существенно упрощает процедуру расчета фрактальной размерности изображения; поскольку одной из задач данного раздела является получение связи между морфологическими характеристиками колонии микроорганизмов и фрактальной размерностью изображения, дискретное представление упрощает решение данной проблемы.
Объект моделирования. В качестве объекта моделирования выбрана колония микроорганизмов одного вида, растущая на плоской твердой основе (питательной среде). Область роста колонии представляет собой ограниченную часть плоскости. Процессы роста колонии и деления клеток происходят на по-
1 Результаты данного раздела опубликованы в наших работах: Арзамасцев А. А., Слетков Д. В. Дискретная модель роста популяции микроорганизмов на плоскости // Вестн. Тамб. ун-та. Сер. Естеств. и техн. науки. – Тамбов, 2005. – Т. 10, вып. 1. – С. 112; Слетков Д. В., Арзамасцев А. А. Дискретная математическая модель формообразования колонии микроорганизмов, растущих на плоскости // Вестн. Тамб. ун-та. Сер. Естеств. и техн. науки. – Тамбов, 2005. – Т. 10, вып. 2. – С. 193-195; Слетков Д. В., Арзамасцев А. А. Морфология и кинетика роста колоний микроорганизмов на плоскости. Результаты вычислительного эксперимента // Вестн. Тамб. ун-та. Сер. Естеств. и техн. науки. – Там-
бов, 2005. – Т. 10, вып. 3. – С. 277-291.
114
верхности среды. Потребление питательного вещества (субстрата) каждой клеткой происходит из среды, расположенной с ней в непосредственной близости. Концентрация субстрата в начале процесса обычно бывает постоянной по всей области. В процессе роста клетки концентрация питательного вещества в области, непосредственно к ней прилегающей, снижаться. Однако со временем может происходить ее восполнение в результате диффузии из соседних областей. Для каждой системы «микроорганизмы – субстрат» существует свой набор кинетических параметров, характеризующих скорости роста и деления клеток, потребления субстрата, отмирания клеток, диффузии субстрата.
Основные допущения модели и их реализуемость.
1. Область распространения микроорганизмов представляет ограниченную часть плоскости с нанесенной дискретной сеткой. Ее можно представить в виде квадратной матрицы, состоящей из ячеек размером N × N (реально используется 100 × 100 ячеек, но при необходимости размер матрицы может быть изменен). Такое представление хорошо соответствует росту реальных колоний микроорганизмов на твердых субстратах.
2.Единицей времени в модели является одна итерация; все временные параметры в модели задаются в количестве итераций. Отсчет времени начинается с нуля итераций.
3.Среда (питательное вещество) распределено по ячейкам области распространения; количество питательного вещества в
начальный момент времени может быть задано как различным для каждой ячейки (задается распределением Q0(i, j), где (i, j) координаты ячейки) так и одинаковым для всех ячеек Q0.
4.Микроорганизм потребляет питательное вещество, которое находится в его ячейке; потреблять питательное вещество из других ячеек он не может. За одну итерацию микроорганизм
потребляет Q– питательного вещества. Такое представление хорошо соответствует реальной ситуации, при которой микроорганизм, растущий на твердом субстрате, потребляет его лишь из близлежащей области.
5.Диффузия питательного вещества в системе задается посредством массопереноса из соседних ячеек, который происходит на каждой итерации. Колонии микроорганизмов могут расти
115

на различных «твердых» средах, каждая из которых имеет различные физико-химические показатели. Таким образом, диффузия в различных средах будет происходить с различной скоростью; этой скорости в модели соответствует коэффициент диффузии η. Диффузионный массоперенос осуществляется согласно уравнениям:
Q1= (Q2 – Q1) η,
Q2= (Q1 – Q2) η,
где Q1 и Q2 – количество питательного вещества перенесенного из ячейки 2 в 1 и из 1 в 2 соответственно. Для каждой ячейки диффузионный массоперенос осуществляется только в четыре (две по вертикали и две по горизонтали) ближайших ячейки (см. рис. 2.44. (1)). Если ячейка находится на границе области распространения или является угловой, то таких ячеек может быть две или три (см. рис. 2.44. (2) и (3)). Это объясняется тем, что для массопереноса необходимо взаимосоприкосновение ячеек, которое достигается лишь в случаях, показанных на рис. 2.44.
Рис. 2.44. Схема массопереноса для различных ячеек области распространения: 1 – для ячейки, находящейся внутри области; 2 – для ячейки, находящейся на границе области; 3 – для угловой ячейки
116
6.Количество питательного вещества в системе (ячейках) может быть восполнено извне, что позволяет моделировать незамкнутую систему, сообщающуюся с внешней средой. Поступ-
ление питательного вещества из внешней среды в одну ячейку за одну итерацию задается распределением Q+(i, j), где (i, j) координаты ячейки в которую поступает питательное вещество. Если данный параметр одинаков для всех ячеек то он обознача-
ется как Q+. Восполнение питательного вещества происходит с первой итерации.
7.Уравнение материального баланса для ячейки (i, j) для итерации t задается уравнением:
Qt (i, j) = Qt–1(i, j) – δ(i, j) Q– + Q+(i, j) + Qtd
где δ(i, j) – функция наличия биологического объекта в ячейке (i, j); она равняется 1, если в ячейке (i, j) имеется такой объект, 0, если его там нет; Qtd – диффузионный массоперенос.
8.Одна клетка занимает одну ячейку области распространения. Пока данная клетка жива, данную ячейку другая клетка занять не может. Клетка неподвижна в течение своей жизни, это объясняется тем, что моделируется выращивание колонии микроорганизмов на «твердых» средах на которых они не могут перемещаться.
9.Изменение формы колонии происходит только за счет размножения клеток. В процессе размножения дочерний микроорганизм занимает одну из 4-х ближайших ячеек, если он располагается по внутренней ячейке, одну из трех или одну из двух ячеек, если он находится на границе области распространения или в угловой ячейке (рис. 2.45). Ячейка может быть занята дочерней клеткой лишь при условии, что в ней нет другого микроорганизма. Поскольку для образования дочерней клетки необходим контакт двух ячеек, процесс размножения возможен только для ячеек соприкасающихся по горизонтали или по вертикали (рис. 2.45). При наличии нескольких свободных ячеек вероятность выбора места дочерней клеткой зависит от количества питательного вещества находящегося в ячейке. Чем больше питательного вещества в ячейке, тем выше вероятность того,
117
что дочерняя клетка замет эту ячейку. Если рядом с родительским микроорганизмом нет свободной ячейки, размножения клетки не происходит. Размножение осуществляется за одну итерацию. Возраст дочерней клетки считается с момента деления. Деление на возрасте родительской клетки не сказывается.
Промежуток времени между делениями микроорганизма задается распределением вероятности P (τd) со средним значением τd. Время следующего деления клетки высчитывается в начальный момент времени и после каждого процесса деления. Если клетка готова к делению, а рядом с ней нет свободных ячеек (т. е. деление клетки на данной итерации невозможно), то на следующей итерации клетка сохраняет способность к делению. Данное допущение соответствует механизму ассиметричного деления, характерного для многих микроорганизмов.
10. Максимальная продолжительность жизни клетки задается распределением вероятности со средним значением τl. Этот параметр определяется при образовании клетки. Клетка может погибнуть и раньше, если на какой-либо итерации количество питательного вещества в его ячейке станет равным нулю.
Рис. 2.45. Положения материнской клетки (1) и возможные расположения дочерних клеток (2)
118
11.Параметры τl, τd характеризуют микроорганизм и среду, на которой выращиваются клетки; для одних и тех же микроорганизмов данные параметры могут различаться на различных средах. Однако для заданной системы «микроорганизмы – среда» будем считать эти параметры постоянными.
12.В начальный момент времени микроорганизмы засеваются в n ячеек области распространения. Их положение может быть как случайным (самопроизвольный засев), так и задаваться координатами клеток, в которых они расположены (искусственный засев). Возраст микроорганизмов находящихся в системе в
начальный момент времени задается распределением со средним значением τ*.
Математическая модель. В начальный момент времени имеется набор клеток:
C ={C(i1 , j1 ) ,C(i2 , j2 ) ,K,C(in , jn )} |
(2.98) |
||
1 |
2 |
n |
|
где C – множество клеток системы, n – общее количество клеток в начальный момент времени, (ik, jk) – индексы k-той клетки.
Для каждой клетки должно выполняться неравенство
l, m Cl, Cm C l≠m |il – im| + |jl – jm| > 0 |
(2.99) |
которое означает, что каждая клетка в системе имеет отдельную ячейку.
Выбор параметров клеток в начальный момент времени осуществляется в соответствии с выражениями:
Ck(ik , jk ) {τkl, τkd, τk, τku} τk= τku=0;
τkl P (τl); τkd P (τd);0 ≤ i ≤ N, 0 ≤ j ≤ N, |
(2.100) |
где τk – возраст k-й клетки, τku – время прошедшее с последнего деления k-й клетки, τkl – максимально возможная продолжительность жизни k-й клетки, определяемая как число из множества случайных чисел принадлежащих распределению P(τl), τkd – время до следующего деления k-й клетки, определяемое как число
119