

Математическое моделирование / Arz_(2010)_Mathematical_&_Computer_Modelling
.pdfРис. 2.52. Зависимость количества живых клеток в популяции от модельного времени для 636 популяций, различающихся начальным положением и возрастом клеток. Значения начальных условий и параметров: Q0 = 100; Q– = 1;
Q+ = 0; η = 0,05; τl = 100; τd = 10
Рис. 2.53. Зависимость фрактальной размерности изображения популяции от модельного времени для 636 популяций, различающихся начальным положением и возрастом клеток. Значения начальных условий и параметров: Q0 = 100;
Q–= 1; Q+ = 0; η = 0,05; τl = 100; τd = 10
130
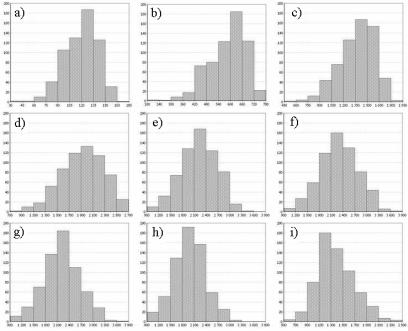
Представляет значительный интерес расчет по данным, приведенным на рис. 2.52 гистограмм распределений численности популяций для различных моментов времени (рис. 2.54). Из этого рисунка видно, что во многих случаях полученные гистограммы отличаются от нормального закона распределения, что указывает на несимметричный характер отклонений от среднего значения. В начальный период роста популяции отклонения наблюдаются в сторону увеличенных значений (рис. 2.54, a) – d)). При средних значениях модельного времени распределения близки к нормальному (рис. 2.54, e) – g)), и в дальнейшем отклонения наблюдаются в сторону уменьшения значений (рис. 2.54, h) – i)).
Рис. 2.54. Гистограммы распределений численности популяций. По оси абсцисс - количество живых клеток в популяции, по оси ординат – количество кинетических кривых, для которых численность популяции попала в данный интервал, для моментов времени: a) – 50; b) – 100; c) – 150; d) – 200; e) – 250; f) – 300; g) – 350; h) – 400 и i) – 500 итераций
131
На рис. 2.55 приведены усредненная кинетическая кривая роста и кривые, ограничивающие область полученных значений численности популяции. Видно, что для экспоненциальной фазы развития средние значения численности популяции очень близки к максимально возможным, в то время как в конце процесса они становятся близкими к минимально возможным (нулевым) значениям. Данное обстоятельство согласуется также с данными, приведенными на рис. 2.54. На рис. 2.55 виден также значительный разброс численности популяции в одинаковые моменты времени, что связано с различиями в начальном положении и возрасте клеток.
На рис. 2.56 отдельно показана усредненная кинетическая кривая роста. На ней можно выделить следующие характерные участки. Участок AB соответствует экспоненциальному росту, который обычно наблюдается в условиях отсутствия каких-либо ограничений. Участок BC соответствует фазе линейного роста, что связано с ограничением питательного вещества. Начало этого участка соответствует также модельному времени 160–170 итераций и образованию пустот внутри развивающихся кластеров (см. далее). Это характерное время практически не зависит от начального расположения и возраста клеток.
Необходимо отметить, линейный рост характерен для популяций, испытывающих в своем развитии ограничения различными факторами, например, температурой, концентрациями веществ и т. д. Интересной особенностью данного вычислительного эксперимента является то, что не вводя в модель подобных ограничений в явном виде, а задав лишь поведение популяции в виде набора простых правил, мы получили аналогичные результаты.
Участок CD является переходным к режиму отмирания популяции – участок DE. Эти особенности также являются характерными для развития большинства популяций.
Кривая, изображенная на рис. 2.56 может быть описана с помощью традиционных детерминированных математических моделей, которые, однако, не могут учесть стохастических особенностей развития популяции, показанных на рис. 2.55.
132

Рис. 2.55. Усредненная кинетическая кривая и огибающие сверху и снизу для семейства кривых, изображенных на рис. 2.52
Рис. 2.56. Усредненная кинетическая кривая для семейства кривых, изображенных на рис. 2.52
133
Влияние морфологических изменений в популяции на кинетику ее роста. Поскольку в предыдущих вычислительных экспериментах мы показали, что существует связь морфологических и кинетических параметров в процессе роста популяции, в данном разделе проведем анализ указанных особенностей на двух характерных примерах (рис. 2.57–2.62).
В первом примере кинетика роста популяции показана на рис. 2.57, ее внешний вид в различные моменты времени на рис.2.58, а фрактальная размерность изображения на рис. 2.59. Отличительной особенностью данного примера является унимодальный характер кинетической кривой (рис.2.57).
Начальное развитие кластеров и их положение показано на рис. 2.58 a). На кинетической кривой (рис. 2.57) этот момент времени соответствует фазе экспоненциального роста. В этот период имеется ярко выраженная зависимость фрактальной размерности изображения от времени (рис. 2.59). На рис. 2.58 b) происходит образование внутренних полостей, в связи, с чем на кинетической кривой (рис. 2.57) наблюдается переход от экспоненциальной фазы к фазе с постоянной скоростью роста. Это связано с тем, что распространение популяции начинает ограничиваться питательным ресурсом. Этот переход сопровождается локальным максимумом фрактальной размерности (рис. 2.59). Фрактальная размерность изображения популяции (рис. 2.59) изменяется только в экспоненциальной фазе развития и практически постоянна в других фазах; из этого можно сделать вывод от том, что фрактальную размерность можно использовать для определения кинетических характеристик популяции начальный период. Следующий этап морфологических изменений популяции показан на рис. 2.58 c). Он связан с достижением максимальной численности и началом процесса схлопывания межкластерных перегородок. Данный процесс сопровождается резким сокращением численности. На рис. 2.58 d) показан момент, когда уже завершилось схлопывание этих перегородок и образовался общий фронт популяции. Из-за движения этого фронта значительное время численность популяции лишь незначительно падает, т. к. идет процесс освоения новых пищевых ресурсов.
134
Во втором примере кинетика роста популяции показана на рис. 2.60, ее внешний вид в различные моменты времени на рис.2.61, а фрактальная размерность изображения на рис. 2.62. Отличительной особенностью данного примера является бимодальный характер кинетической кривой (рис. 2.60).
Начальное развитие кластеров и их положение показано на рис. 2.61 a). На кинетической кривой (рис. 2.60) этот момент времени соответствует фазе экспоненциального роста. В этот период также имеется ярко выраженная зависимость фрактальной размерности изображения от времени (рис. 2.62). На рис. 2.61 b) происходит образование внутренних полостей, в связи, с чем на кинетической кривой (рис. 2.60) наблюдается переход от экспоненциальной фазы к фазе с постоянной скоростью роста. Однако в отличие от рис. 2.57 данный процесс выражен слабее из-за того, что образование полостей совпало с началом взаимодействия соседних кластеров. Этот переход сопровождается также локальным максимумом фрактальной размерности (рис. 2.62). Следующий этап морфологических изменений популяции показан на рис. 2.61 c). Он связан с достижением максимальной численности и началом процесса схлопывания межкластерных перегородок. Данный процесс сопровождается сокращением численности, вслед за которым наблюдается ее рост, связанный с образованием единого фронта (рис. 2.61 d)). На рис. 2.61 e) показано изображение популяции в момент достижения второго локального максимума на кинетической кривой (рис. 2.60). Дальнейшее резкое уменьшение численности связано с соприкосновением двух фронтов (рис. 2.61 f)).
Обобщение морфологических характеристик и соответствующих им особенностей на кинетических кривых показано в табл. 2.10.
135
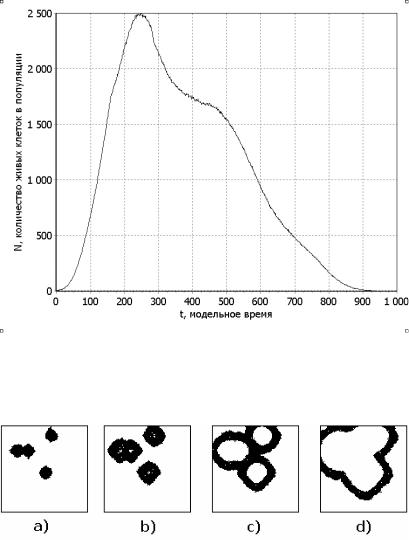
Рис. 2.57. Зависимость количества живых клеток от модельного времени для одной из популяций, показанных на рис. 2.52. Пример 1
Рис. 2.58. Внешний вид популяции, кинетика роста которой изображена на рис. 2.57, в моменты времени: a) – 100, b) – 160, c) – 250, d) – 350 итераций
136

Рис. 2.59. Зависимость фрактальной размерности изображения от модельного времени для популяции, кинетика роста которой изображена на рис. 2.57
Рис. 2.60. Зависимость количества живых клеток от модельного времени для одной из популяций, показанных на рис. 2.52. Пример 2
137

Рис. 2.61. Внешний вид популяции, кинетика роста которой изображена на рис. 2.60, в моменты времени: a) – 100, b) – 160, c) – 220, d) – 350, e) – 450, f) – 500 итераций
Рис. 2.62. Зависимость фрактальной размерности изображения от модельного времени для популяции, кинетика роста которой изображена на рис. 2.60
138
Таблица 2.10
Морфологические изменения в популяции и соответствующие им особенности на кинетических кривых роста
Морфологические изменения |
Особенность на кинетической |
|||
кривой |
||||
|
|
|
||
Образование |
пустоты |
внутри |
Переход от экспоненциальной фазы к |
|
развивающихся кластеров |
попу- |
фазе развития с постоянной скоростью |
||
ляции |
|
|
роста |
|
Разрушение больших фронтов |
Резкое падение численности популя- |
|||
|
|
|
ции |
|
Разрушение |
перегородок |
между |
Падение численности популяции сразу |
|
отдельными кластерами |
|
после достижения максимального зна- |
||
|
|
|
чения |
|
Движение популяционного фрон- |
Практически постоянная на длитель- |
|||
та по части плоскости, где имеет- |
ном временном интервале (слабо убы- |
|||
ся достаточное количество пита- |
вающая или слабо возрастающая) чис- |
|||
тельного вещества |
|
ленность популяции |
||
Момент начального соприкосно- |
Максимум численности популяции |
|||
вения развивающихся кластеров |
|
|||
популяции |
|
|
|
|
Бифуркации в процессе роста популяций на плоскости.
В ходе вычислительных экспериментов была обнаружена следующая интересная особенность. Оказалось, что при фиксированном начальном расположении клеток (в вершинах квадрата) наблюдалось разделение кинетических кривых на четко выраженные семейства (рис. 2.63), что позволило говорить о наличие бифуркации в системе.
На рис. 2.63 представлено семейство кинетических кривых, а на рис. 2.64 семейство зависимостей фрактальной размерности изображения от модельного времени, которые представляют собой результаты 100 вычислительных экспериментов со следующими значениями параметров и начальных условий: начальное количество питательного вещества – 100; скорость потребления питательного вещества – 1; скорость восполнения питательного вещества – 0; коэффициент диффузии – 0,05; среднее значение максимальной продолжительности жизни – 100; среднее значение времени между делениями микроорганизма – 10. Начальное положение клеток – 4 ячейки с координа-
139