

3. Восстановление объёма крови при обезвоживании организма
Уменьшение общего объёма жидкости, например в результате кровопотери, при обильной рвоте, диарее вызывает высвобождение ренина. Этому способствует также снижение импульсации от барорецепторов предсердий и артерий в результате уменьшения внутрисосудистого объёма жидкости. В результате увеличивается продукция ангиотензина II, наиболее мощного стимулятора секреции альдостерона. Повышение концентрации альдостерона в крови вызывает задержку ионов натрия, что является сигналом для осморецепторов гипоталамуса и секреции из нервных окончаний передней доли гипофиза АДГ, стимулирующего реабсорбцию воды из собирательных трубочек. Ангиотензин II, оказывая сильное сосудосуживающее действие, повышает АД и, кроме этого, усиливает жажду. Поступающая с питьём вода в большей мере, чем это происходит в норме, задерживается в организме. Увеличение объёма жидкости а, также повышение АД приводят к устранению стимула, который вызвал активацию ренинангиотензиновой системы, секрецию альдостерона и восстановление объёма крови (рис. 11-35).
4. Гиперальдостеронтм
Гиперальдостеронизм - заболевание, вызванное гиперсекрецией альдостерона надпочечниками. Причиной первичного гиперальдостеронизма (синдром Кона) примерно у 80% больных является аденома надпочечников, в остальных случаях - диффузная гипертрофия клеток клубочковой зоны, вырабатывающих альдостерон. При первичном гиперальдостеронизме избыток альдостерона усиливает реабсорбцию натрия в почечных канальцах. Увеличение концентрации Na+ в плазме служит стимулом к секреции АДГ и задержке воды почками. Кроме того, усиливается выведение ионов калия, магния и протонов. В результате развиваются гипернатриемия, вызывающая, в частности, гипертонию, гипер-волемию и отёки, а также гипокалиемия, ведущая к мышечной слабости, возникают дефицит магния и лёгкий метаболический алкалоз.
Вторичный гиперальдостеронизм встречается гораздо чаще, чем первичный, и может быть связан с рядом состояний (например, сердечная недостаточность, хронические заболевания почек, а также сопровождающиеся нарушением кровоснабжения опухоли, секретирующие ренин). При вторичном гиперальдостеронизме у больных наблюдают повышенный уровень ренина и ангиотензина II, что стимулирует кору надпочечников продуцировать и секретировать избыточное количество альдостерона. Клинические симптомы менее выражены, чем при первичном альдостеронизе. Одновременное определение концентрации альдостерона и активности ренина в плазме позволяет окончательно дифференцировать первичный (активность ренина в плазме снижена) и вторичный (активность ренина в плазме повышена) гиперальдостеронизм.
В. Предсердный натриуретический фактор (ПНФ)
Это пептид, содержащий 28 аминокислот с единственным диеульфидным мостиком. ПНФ синтезируется, главным образом, в кардиомио-цитах предсердий, и хранится в виде препро-гормона, состоящего из 126 аминокислотных остатков.
602
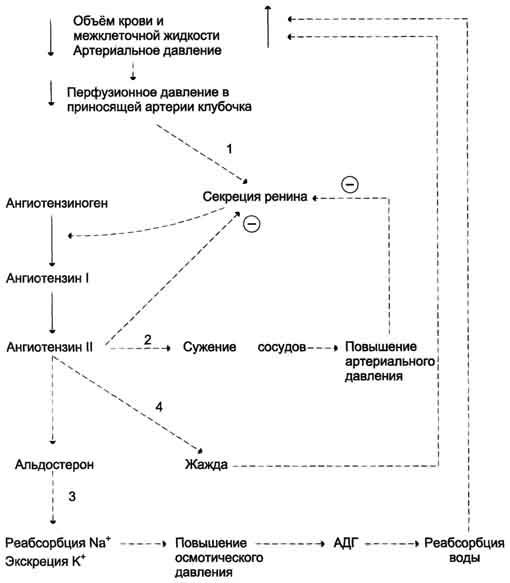
Рис. 11-35. Схема восстановления объёма крови при кровопотере и обезвоживании организма. 1 - уменьшение объёма жидкости и снижение АД активируют систему ренинангиотензинальдостерон; 2 - ангиотензин II вызывает сужение сосудов, что является экстренной мерой для поддержания АД; 3 - альдостерон стимулирует задержку натрия, вследствие чего происходит высвобождение вазопрессина и усиливается реабсорбция воды; 4 - ангиотензин II вызывает также чувство жажды, что способствует увеличению жидкости в организме.
Основным фактором, регулирующим секрецию предсердного натрийуретического фактора, является увеличение АД. Другие стимулы секреции - увеличение осмолярности плазмы, повышение частоты сердцебиений, повышенный уровень катехоламинов и глюкокортикоидов в крови.
Основные клетки-мишени ПНФ - почки, периферические артерии. В почках ПНФ стимулирует расширение приносящих артериол, усиление почечного кровотока, увеличение скорости фильтрации и экскреции ионов натрия. В периферических артериях ПНФ снижает тонус гладких мышц и соответственно расширяет артериолы (рис. 11-36). Таким образом, суммарным действием ПНФ является увеличение экскреции Na+ и понижение АД.
Механизм передачи сигнала ПНФ не включает актвивацию G-белка. Рецептор ПНФ имеет доменное строение: домен связывания с лигандом,
603
локализованный во внеклеточном пространстве, и один домен, пронизывающий мембрану и обладающий активностью гуанилатциклазы. В отсутствие ПНФ его рецептор находится в фосфорилированном состоянии и неактивен. Связывание ПНФ с рецептором вызывает кон-формационные изменения и возрастание гуанилатциклазной активности рецептора. В результате ГТФ превращается в циклический ГМФ (цГМФ), который активирует протеинкиназу G (см. раздел 5).
ПНФ обычно рассматривают как физиологический антагонист ангиотензина II, поскольку под его влиянием возникают не сужение просвета сосудов и задержка натрия, а, наоборот, расширение сосудов и увеличение почечной экскреции соли.
87. Регуляция обмена ионов кальция и фосфатов. Строение, биосинтез и механизм действия паратгормона, кальцитонина и кальцитриола. Причины и проявления рахита, гипо- и гиперпаратиреодизма.
В организме взрослого человека содержится в среднем 1000 г кальция. Основным депо кальция в организме (99% всего кальция от общей массы) являются кости. В костях около 99% кальция присутствует в малорастворимой форме кристаллов гидроксиапатита [Са10(РО4)6(ОН)2Н2О]. В виде фосфатных солей в костях находится лишь 1% кальция, который может легко обмениваться и играть роль буфера при изменениях концентрации кальция в плазме крови. Другой фонд кальция (1% от общей массы кальция) - кальций плазмы крови. В плазму крови кальций поступает из кишечника (с водой и пищей) и из костной ткани (в процессе резорбции).
Кальций - не только структурный компонент костной ткани. Ионы кальция играют ключевую роль в мышечном сокращении, увеличивают проницаемость мембраны клеток для ионов калия, влияют на натриевую проводимость клеток, на работу ионных насосов, способствуют секреции гормонов, участвуют в каскадном механизме свёртывания крови. Кроме этого, они служат важнейшими посредниками во внутриклеточной передаче сигналов.
Концентрация кальция внутри клеток зависит от его концентрации во внеклеточной жидкости. Пределы колебаний общей концентрации Са2+ в плазме крови здоровых людей составляют
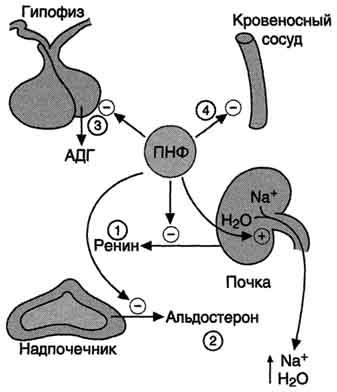
Рис. 11-36. Биологическое действие ПНФ. 1 - ингибирует выделение ренина; 2 - ингибирует секрецию альдостерона; 3 - ингибирует секрецию АДГ; 4 - вызывает релаксацию сосудов.
2,12-2,6 ммоль/л, или 9-11 мг/дл. Кальций плазмы крови представлен в виде:
несвязанного, ионизированного кальция (около 50%);
ионов кальция, соединённых с белками, главным образом, с альбумином (45%);
недиссоциирующих комплексов с цитратом, сульфатом, фосфатом и карбонатом (5%).
Биологически активной фракцией является ионизированный кальций, концентрация которого поддерживается в пределах 1,1-1,3 ммоль/л.
Изменение уровня кальция может привести к нарушению многих процессов: изменению порога возбудимости нервных и мышечных клеток, нарушению функционирования кальциевого насоса, снижению активности ферментов и нарушению гормональной регуляции метаболизма. Концентрация Са2+ в плазме регулируется с высокой точностью: изменение её всего на 1% приводит в действие гомеостатические механизмы, восстанавливающие равновесие.
604
Основными регуляторами обмена Са2+ в крови являются паратгормон, калыщтриол и каль-цитонин.
А. Паратгормон
Паратгормон (ПТГ) - одноцепочечный полипептид, состоящий из 84 аминокислотных остатков (около 9,5 кД), действие которого направлено на повышение концентрации ионов кальция и снижение концентрации фосфатов в плазме крови.