

74. Непрямое дезаминирование аминокислот. Схема процесса, субстраты, ферменты, кофакторы.
![]()

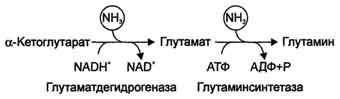
Непрямое дезаминирование аминокислот происходит при участии 2 ферментов: аминотрансферазы (кофермент ПФ) и глутаматдегидрогеназы (кофермент NAD+).
Биологическая роль непрямого дезаминирования. А - при катаболизме почти все природные аминокислоты сначала передают аминогруппу на а-кетоглутарат в реакции трансаминирования с образованием глутамата и соответствующей кетокислоты. Затем глутамат подвергается прямому окислительному дезаминированию под действием глутаматдегидрогена-зы, в результате чего получаются а-кетоглутарат и аммиак; Б - при необходимости синтеза аминокислот и наличии необходимых а-кетокислот обе стадии непрямого дезаминирования протекают в обратном направлении. В результате восстановительного аминирования а-кетоглутарата образуется глутамат, который вступает в трансаминирование с соответствующей а-кетокислотой, что приводит к синтезу новой аминокислоты.
75.
Основные источники аммиака в организме
человека. Токсичность аммиака. Роль
глутамина и аспарагина в обезвреживании
аммиака. Глутаминаза почек, образование
и выведение солей аммония.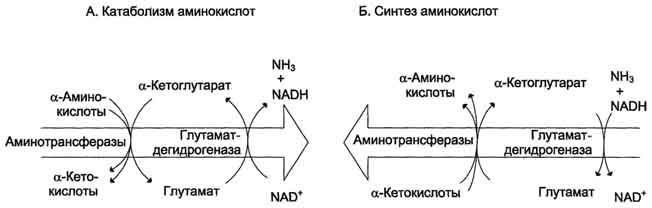
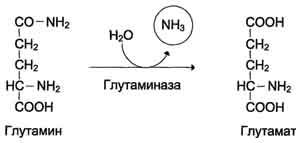
ужат мышцы, мозг и печень. С током крови глутамин транспортируется в кишечник и почки.
В клетках кишечника под действием фермента глутаминазы происходит гидролитическое освобождение амидного азота в виде аммиака:
Образовавшийся в реакции глутамат подвергается трансаминированию с пируватом. ос-Аминогруппа глутаминовой кислоты переносится в состав аланина (рис. 9-10). Большие количества аланина поступают из кишечника в кровь воротной вены и поглощаются печенью. Около 5% образовавшегося аммиака удаляется в составе фекалий, небольшая часть через воротную вену попадает в печень, остальные ~90% выводятся почками.
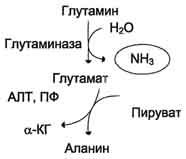
Рис. 9-10. Метаболизм азота глутамина в кишечнике.
В почках также происходит гидролиз глутамина под действием глутаминазы с образованием аммиака. Этот процесс является одним из механизмов регуляции кислотно щелочного равновесия в организме и сохранения важнейших катионов для поддержания осмотического давления. Глутаминаза почек значительно индуцируется при ацидозе, образующийся аммиак нейтрализует кислые продукты обмена и в виде аммонийных солей экскретируется с мочой (рис. 9-11). Эта реакция защищает организм от излишней потери ионов Na+ и К+, которые также могут использоваться для выведения анионов и утрачиваться. При алкалозе количество глутаминазы в почках снижается.
В почках образуется и выводится около 0,5 г солей аммония в сутки.
Высокий уровень глутамина в крови и лёгкость его поступления в клетки обусловливают использование глутамина во многих анаболических процессах. Глутамин - основной донор азота в организме. Амидный азот глутамина используется для синтеза пуриновых и пиримидиновых
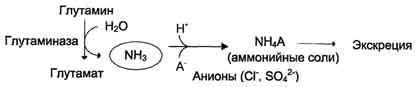
Рис. 9-11. Метаболизм амидного азота глутамина в почках.
479
нуклеотидов, аспарагина, аминосахаров и других соединений (рис. 9-12).
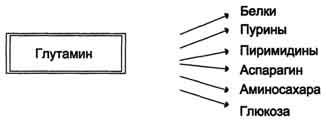
Рис. 9-12. Пути использования глутамина в организме.
Ещё одной реакцией обезвреживания аммиака в тканях можно считать синтез аспарагина под действием аспарагинсинтетазы.
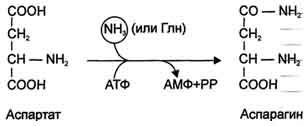
Существуют 2 изоформы этого фермента - глутаминзависимая и аммиакзависимая, которые используют разные доноры амидных групп. Первая функционирует в животных клетках, вторая преобладает в бактериальных клетках, но присутствует и у животных. Однако такой путь обезвреживания аммиака в клетках человека используется редко и к тому же требует больших энергетических затрат (энергию двух макроэргических связей), чем синтез глутамина.
Наиболее значительные количества аммиака обезвреживаются в печени путём синтеза мочевины. В первой реакции процесса аммиак связывается с диоксидом углерода с образованием карбамоилфосфата, при этом затрачиваются 2 молекулы АТФ. Реакция происходит в митохондриях гепатоцитов под действием фермента карбамоилфос-фатсинтетазы I. Карбамоилфосфатсинтетаза II локализована в цитозоле клеток всех тканей и участвует в синтезе гшримидиновых нуклеотидов (см. раздел 10). Карбамоилфосфат затем включается в орнитиновый цикл и используется для синтеза мочевины.
В мозге и некоторых других органах может протекать восстановительное аминирование α-кетоглутарата под действием глутаматдегидрогеназы, катализирующей обратимую реакцию. Однако этот путь обезвреживания аммиака в тканях используется слабо, так как глутаматдегидрогеназа катализирует преимущественно реакцию дезаминирования глутамата. Хотя, если учитывать последующее образование глутамина, реакция выгодна для клеток, так как способствует связыванию сразу 2 молекул NH3.
Из мышц и кишечника избыток аммиака выводится преимущественно в виде аланина. Этот механизм необходим, так как активность глутаматдегидрогеназы в мышцах невелика и непрямое дезаминирование аминокислот малоэффективно. Поэтому в мышцах существует ещё один путь выведения азота. Образование аланина в этих органах можно представить следующей схемой (см. схему ниже).
Аминогруппы разных аминокислот посредством реакций трансаминирования переносятся на пируват, основным источником которого служит процесс окисления глюкозы.
Мышцы выделяют особенно много аланина в силу их большой массы, активного потребления
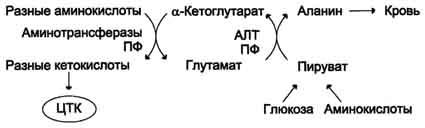
Схема

480
глюкозы при физической работе, а также потому, что часть энергии они получают за счёт распада аминокислот. Образовавшийся аланин поступает в печень, где подвергается непрямому дезаминированию. Выделившийся аммиак обезвреживается, а пируват включается в глюконеогенез. Глюкоза из печени поступает в ткани и там, в процессе гликолиза, опять окисляется до пирувата (рис. 9-13).
Образование аланина в мышцах, его перенос в печень и перенос глюкозы, синтезированной в печени, обратно в мышцы составляют глюкозо-аланиновый цикл, работа которого сопряжена с работой глюкозо-лактатного цикла (см. раздел 7).
Совокупность основных процессов обмена аммиака в организме представлена на рис. 9-14. Доминирующими ферментами в обмене аммиака служат глутаматдегидрогеиаза и глутаминсинтетаза.
76. Оринитиновый цикл мочевинообразования. Химизм, место протекания процесса. Энергетический эффект процесса, его регуляция. Возрастные особенности использования аммиака в детском возрасте. Количественное определение мочевины сыворотки крови, клиническое значение.
Окислительное
дезаминирование глутамата происходит
в митохондриях. Ферменты орнитинового
цикла распределены между митохондриями
и цитозолем. Поэтому необходим
трансмембранный перенос глутамата,
цитруллина и орнитина с помощью
специфических транслоказ. На схеме
показаны пути включения азота двух
разных аминокислот (аминокислота 1 и
аминокислота 2) в молекулу мочевины: •
одна аминогруппа - в виде аммиака в
матриксе митохондрии; • вторую аминогруппу
поставляет аспартат цитозоля.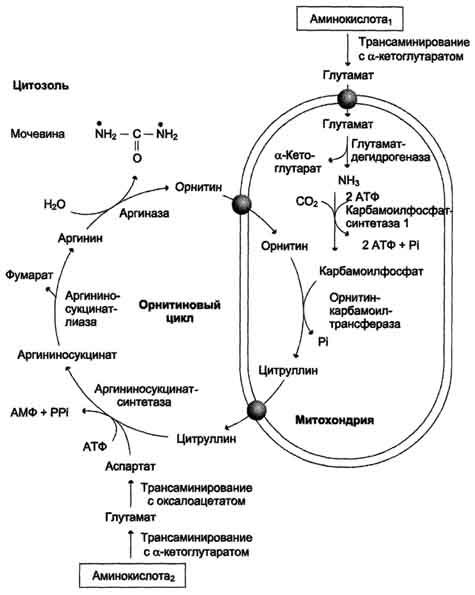
Энергетический баланс процесса
В реакциях орнитинового цикла расходуются четыре макроэргических связи трёх молекул АТФ на каждый оборот цикла. Однако процесс превращения аминокислот в безазотистые остатки и мочевину имеет пути компенсации энергозатрат:
при включении фумарата в ЦТК на стадии дегидрирования малата образуется NADH, который обеспечивает синтез 3 молекул АТФ;
при окислительном дезаминировании глу-тамата в разных органах также образуется NADH, соответственно - ещё 3 молекулы АТФ.
Количественное определение мочевины в сыворотке крови.
Принцип метода. Диацетилмонооксим в кислой среде и в присутствии тиосемикарбазида и ионов трехвалентного железа образует с мочевиной красный комплекс.
Техника выполнения.
Смешать реактивы, на 10 минут поместить в кипящую водяную баню все три пробирки, предварительно закрыв их отверстие фольгой. Затем быстро охлаждают все пробирки водой и колориметрируют при длине волны 490-540 нм опытную и калибровочную пробы против контроля в кюветах толщиной 0,5 см. Формула для расчета:
Аоп./Аэ×16,6 (ммоль/л).
Норма содержания мочевины в крови: 2,5-8,3 ммоль/л. Уровень мочевины в крови характеризует выделительную функцию почек.
Аммиак выделяется в моче в виде солей серной и фосфорной кислот. За счет аммиака у детей выделяется относительно больше азота, чем у взрослых. Избыток аммиака в детской моче зависит от неполного превращения его в мочевину. Аммиак входит в состав солей серной и фосфорной кислот, образующихся при расщеплении белка и фосфорсодержащих органических соединений. У взрослого это осуществляется отчасти за счет щелочных земель (Na, К, Са, Mg), поступающих в достаточном количестве с пищей. Детский организм эти соли использует для пластических целей; кроме того, всасывание их в кишечнике несколько затруднено образованием мыл вследствие относительно большого содержания жира в пище ребенка. Повышенное содержание аммиака в моче не говорит об ацидозе и ацидурии, а скорее об алкалопении, указывая на некоторый недостаток щелочей. У старших детей количество аммиака в моче зависит от характера пищи, главным образом от характера ее зольного остатка; при большом количестве овощей поступает много щелочей и, следовательно, меньше выделяется аммиака с мочой; при мясной пище, наоборот, больше образуется кислых продуктов межуточного обмена, нейтрализуемых аммиаком и выделяющихся с мочой в виде соответствующих соединений.
77. Декрбоксилирование АК. Биогенные амины: гистамин, серотонин, ГАМК. Реакции их образования, ферменты и коферменты. Биороль биогенных аминов. Дезаминирование и метилирование аминов как пути из обезвреживания.
Декарбоксилирование аминокислот и их производных
Некоторые аминокислоты и их производные могут подвергаться декарбоксилированию - отщеплению ос-карбоксильной группы. В тканях млекопитающих декарбоксилированию может подвергаться целый ряд аминокислот или их производных: Три, Тир, Вал, Гис, Глу, Цис, Apr, Орнитин, SAM, ДОФА, 5-окситриптофан и др. Продуктами реакции являются СО2 и амины, которые оказывают выраженное биологическое действие на организм (биогенные амины):

Реакции декарбоксилирования необратимы и катализируются ферментами декарбоксилазами. Простетическая группа декарбоксилаз в клетках животных - пиридоксальфосфат. Некоторые декарбоксилазы микроорганизмов могут содержать вместо ПФ остаток пирувата - гистидиндекарбоксилаза Micrococcus и Lactobacilus, SAM-декарбоксилаза Е. coli и др. Механизм реакции напоминает
512
реакцию трансаминирования с участием пиридоксальфосфата и также осуществляется путём формирования шиффова основания ПФ и аминокислоты на первой стадии.
Амины, образовавшиеся при декарбоксилировании аминокислот, часто являются биологически активными веществами. Они выполняют функцию нейромедиаторов (серотонин, дофамин, ГАМК и др.), гормонов (норадреналин, адреналин), регуляторных факторов местного действия (гистамин, карнозин, спермин и др.).