


Фізіологія сільськогосподарських тварин
ВЛАСТИВОСТІ ГЛАДЕНЬКИХ М’ЯЗІВ
Збудливість гладеньких м’язів значно нижча, ніж скелетних.
У гладенькому м’язі ссавців збудження проводиться дуже повільно (від 1 см/с у кишках, до 18 см/с у сечоводі). Воно передається від одного гладенького м’яза до іншого.
Основна відмінність гладеньких м’язів від скелетних. — повільність їх скорочення. Латентний період скорочення гладенького м’яза у 300 разів перевищує скритий період скелетного. Тривалість скорочення в шлунку кроля досягає 5, а в шлунку жаби — 60 с і більше.
Багато гладеньких м’язів скорочується під впливом імпульсів, що виникають у м’язових клітинах, тобто володіють автоматизмом.
На відміну від скелетних, гладенькі м’язи збуджуються деякими хімічними речовинами, що надходять у кров. Так, ацетилхолін стимулює, адреналін гальмує їх активність.
Гладенькі м’язи здатні довго перебувати у тонічному стані при надмірно малій витраті енергії. Це особливо яскраво виявляється у сфінктерах шлунка, жовчного міхура, матки та інших органів. М’язова частина стінки кровоносних судин тварини залишається в тонусі протягом усього її життя.
Добре виражена пластичність гладеньких м’язів має важливе біологічне значення для порожнинних органів, що виконують функцію резервуара. Завдяки пластичності при наповненні шлунка, жовчного міхура, кишок, сечового міхура тиск майже не змінюється.
ФІЗІОЛОГІЯ НЕРВІВ
Відростки нервової клітини, які складаються з осьового циліндра та оболонок, створюють нервові; волокна. Вони можуть бути м’якотні та безм’якотні. У м’якотних, або мієлінових, волокнах зовні є шваннівська оболонка,; утворена шваннівськими клітинами (клітинамисателітами), за якою йде мієлінова оболонка. По ходу волокна через кожні 1–2,5 мм остання переривається, створюючи перехвати Ранв’є. Осьовий циліндр нервового волокна складається з окремих мітохондрій, мікросом та багатьох нейрофібрил, що є продовженням фібрилярної сітки нервової клітини. Спинномозкові та парасимпатичні
442

Розділ 12. Фізіологія м’язів та нервів
нерви складаються з м’якотних волокон. Постгангліонарні волокна симпатичної нервової системи не мають мієлінової оболонки.
Нервові волокна, що проводять імпульси від периферії до нервових центрів, називаються сенсорними, чутливими, доцентровими або аферентними. Інші волокна, що проводять імпульси від центра до периферії, є руховими, моторними, відцентровими, або еферентними.
В організмі тварини нерви звичайно змішані, мають у своєму складі аферентні та еферентні волокна.
Властивості нервових волокон
Збудливість. Нервові волокна володіють збудливістю. Особливо висока вона у м’якотних волокон, що підтверджується даними хронаксії: у моторних волокон ссавців вона становить 0,05–0,2, у симпатичних — до 5 мс.
Провідність. Крім збудливості, основною особливістю нерва є його провідність — здатність проводити збудження. Воно проводиться з допомогою кругових струмів по поверхні нервового волокна від одного перехвату Ранв’є до другого. Мієлінова оболонка виконує конденсаторну та трофічну функції.
У безм’якотних нервових волокнах імпульси поширюються безперервно уздовж тонкої шваннівської оболонки від однієї ділянки збудження до іншої.
На всьому протягу нерва ніякого ослаблення або зменшення швидкості проведення збудження не відбувається. Таке проведення збудження без втрати напруження і швидкості поширення потенціалу називається проведенням без декремента (спаду). Воно властиве всім хребетним тваринам.
Найбільша швидкість проведення збудження у рухових нервах теплокровних — до 160 м/с, у чутливих нервах вона становить до 50 м/с, найменша вона у безм’якотних постгангліонарних симпатичних нервах (0,3–1,5 м/с).
Ізольованість проведення збудження. Периферичні нерви у своєму складі мають тисячі різноманітних нервових волокон (чутливих, рухових, вегетативних), спрямовані до багатьох органів. Так, трійчастий нерв іннервує жувальні м’язи, шкіру голови, очі, носову порожнину, передню частину язика та зуби; сідничний — усі кістки, м’язовий, апарат, судини та шкіру задньої кінцівки. Отже, нормальна
443

Фізіологія сільськогосподарських тварин
діяльність органів можлива у випадку ізольованого проведення імпульсів по кожному волокну зокрема. В протилежному випадку збудження одного волокна викликало б відповідну реакцію з боку багатьох периферичних утворень.
Значення цілісності нерва. По нервовому волокну збудження проводиться лише за умови його анатомічної цілісності та нормального фізіологічного стану.
Осьовий циліндр центрального кінця перерізаного нерва може вростати в його периферичну частину, досягаючи органа, що іннервує. Процес регенерації відбувається досить повільно (3 мм на добу) і в основному залежить від тісного контакту перерізаних частин нерва.
Двобічне проведення збудження. Нервове волокно проводить збудження в обидва боки від місця подразнення. Ця властивість нервового волокна експериментально була доведена російським фізіологом О. І. Бабухіним (1877) на нерві електричного органа нільського сома.
Двобічність проведення збудження по нерву переконливо підтверджується реєстрацією біострумів по обидва боки від місця подразнення.
Функціональна рухомість (лабільність) спинномозкових нервів є найвищою у порівняльно-фізіологічному аспекті. Якщо нервова клітина може відтворити не більше 100 імпульсів за 1 с, то для м’якотного нервового волокна максимальний ритм збудження дорівнює 300–500 імпульсів за 1 с. Висока функціональна рухомість аферентних та еферентних шляхів забезпечує передачу будь-яких ритмів збудження, що виникають у рецепторах без їх трансформації.
Обмін речовин у нерві. Порівняно з іншими органами інтенсивність обміну речовин у нерві дуже низька. Установлено, що сідничний нерв жаби в стані спокою виділяє 0,008 мг вугільної кислоти за 1 хв. Під час роботи нерва споживання кисню та виділення вуглекислоти підвищується на 16–35%.
На відміну від м’яза, нерв не здатний ресинтезувати гексозофосфат з молочної кислоти. Іншою відмінністю обміну речовин нерва та м’яза є утворення аміаку, що вказує на розпад азотистих речовин.
Теплопродукція нерва також дуже мала. При передачі імпульсу по нервовому волокну на відстань 1 см виділяється 10–12 кал. Нерв при збудженні витрачає у 400 000–1 000 000 раз менше енергії, ніж м’яз.
444

Розділ 12. Фізіологія м’язів та нервів
Стомлення нерва. У зв’язку з малими енергетичними затратами і великою інтенсивністю відновних процесів нервове волокно практично невтомне. Це доведено М. Є. Введенським (1884) на нервово- м’язовому препараті жаби. Подразнюючи сідничний нерв жаби протягом 9–10 год, він не виявив ознак втоми у ньому.
Іншим доказом невтомності нерва є тривале прослуховування у ньому за допомогою телефону струмів дії.
Однак без кисню нерв швидко стомлюється і втрачає здатність проводити збудження. Це свідчить про те, що діяльність нерва тісно пов’язана з його окислювальними процесами.
СИНАПСИ
Місця контактів одного нейрона з іншим, або аксона (нейриту) з робочим органом називаються синапсами (грец. synapsis — з’єднання). На одному нейроні кількість синапсів може досягати декількох тисяч. Залежно від місця розміщення синапси ділять на центральні (міжнейронні), периферичні (нервово-м’язові) та ін.
Синаптичні утворення, або бляшки, являють собою своєрідні потовщення розгалужень нейриту з мітохондріями, синаптичними міхурцями та іншими утвореннями. За допомогою електронної мікроскопії в синапсах розпізнають пресинаптйчну та постсинаптичну мембрани та синаптичну щілину шириною близько 200 c , заповнену тканинною рідиною.
Вітчизняними та зарубіжними авторами установлено, що механізм синаптичної передачі збудження тісно пов’язаний з біострумами та хімічними речовинами — медіаторами.
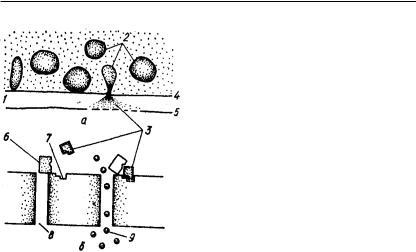
Передавачем збудження у міжнейронних та нервово-м’язових синапсах є ацетилхолін. Закінчення постгангліонарних симпатичних нервових волокон у процесі збудження виділяють норадреналін та незначну кількість адреналіну, складаючих разом симпатин. Імпульс, який надходить до синапсу, звільнює з міхурців медіатор — ацетилхолін. Останній, діючи на постсинаптичну мембрану, підвищує її проникність. Згідно з уявленнями Екклса (1963), молекули медіатору, прикріплюючись до рецепторних ділянок, відкривають пори у мембрані для проходження іонів (рис. 130). У зв’язку з високою концентрацією
445
Фізіологія сільськогосподарських тварин
Рис. 130. Модель синаптичної передачі (за Мільнером):
а — вихід медіатору з синаптичних міхурців через пресинаптичну мембрану (молекули медіатору, сполучаючись з постсинаптичною мембраною, відкривають у ній пори); б — збільшене зображення постсинаптичної мембрани (молекули медіатору прикріплюються до рецепторних ділянок мембрани і зрушують сусідні «пробки», відкриваючи пори у мембрані); 1 — синаптична щілина; 2 — си-
наптичні міхурці; 3 — молекули медіатору; 4 — пресинаптична мембрана; 5 — постсинаптична мембрана; 6 — «пробка» пори; 7 — рецепторна ділянка; 8 — пора постсинаптичної мембрани; 9 — іони, що входять у клітину
натрію у міжклітинній рідині він притягується у клітину негативним зарядом цитоплазми і зменшує внутрішній потенціал приблизно на 0,5 мВ. При цьому виникають кругові електричці струми, що збуджують клітину.
Ацетилхолін, що виділяється, викликає тільки один імпульс, тому що під дією ферментів холінестерази та ацетилхолінестерази він дуже швидко, протягом тисячних долей секунди, розпадається на холін та оцтову кислоту. Після його розпаду мембрана відразу стає непроникною для натрію і за 3–4 мс відновлюється попередній потенціал.
У міжнейронних з’єднаннях виникненню імпульсів передують електричні заряди, які можуть бути збуджуючими або гальмівними постсинаптичними потенціалами (відповідно ЗПСП і ГПСП).
Передача імпульсів через синапси має свої особливості. На відміну від нервового волокна синапси проводять збудження в одному напрямку.
При подразненні рухового нерва м’яз скорочується, але при подразненні м’яза імпульси на руховий нерв не переходять. Однобічність проведення збудження пояснюється тим, що медіатори виділяються лише нервовими закінченнями. Імпульси проходять через синапси з уповільненням. Затримка ця становить близько 1 мс і зумовлена особливостями
446