

Полезные материалы за все 6 курсов / Ответы к занятиям, экзаменам / Биохимия в рисунках и схемах
.pdfРАЗДЕЛ 4. ОБМЕН УГЛЕВОДОВ
УГЛЕВОДЫ – это альдегидо- и кетопроизводные
многоатомных спиртов (альдозы и кетозы) |
|
|
Функции углеводов: |
|
|
энергетическая (моносахариды, гомополисахариды) |
R |
R |
структурная (гетерополисахариды) |
альдоза |
кетоза |
углеводы входят в состав сложных белков – гликопротеинов |
|
|
(ферментов, гормонов, рецепторов, иммуноглобулинов и т.д.) |
|
|
углеводы используются в синтезе нуклеотидов (в синтезе ДНК, РНК, коферментов)
Моносахариды: |
Дисахариды: |
глюкоза |
мальтоза |
фруктоза |
сахароза |
галактоза |
лактоза |
Полисахариды:
крахмал (амилоза и амилопектин) гликоген 






 целлюлоза
целлюлоза 
глюкоза |
галактоза |
фруктоза |
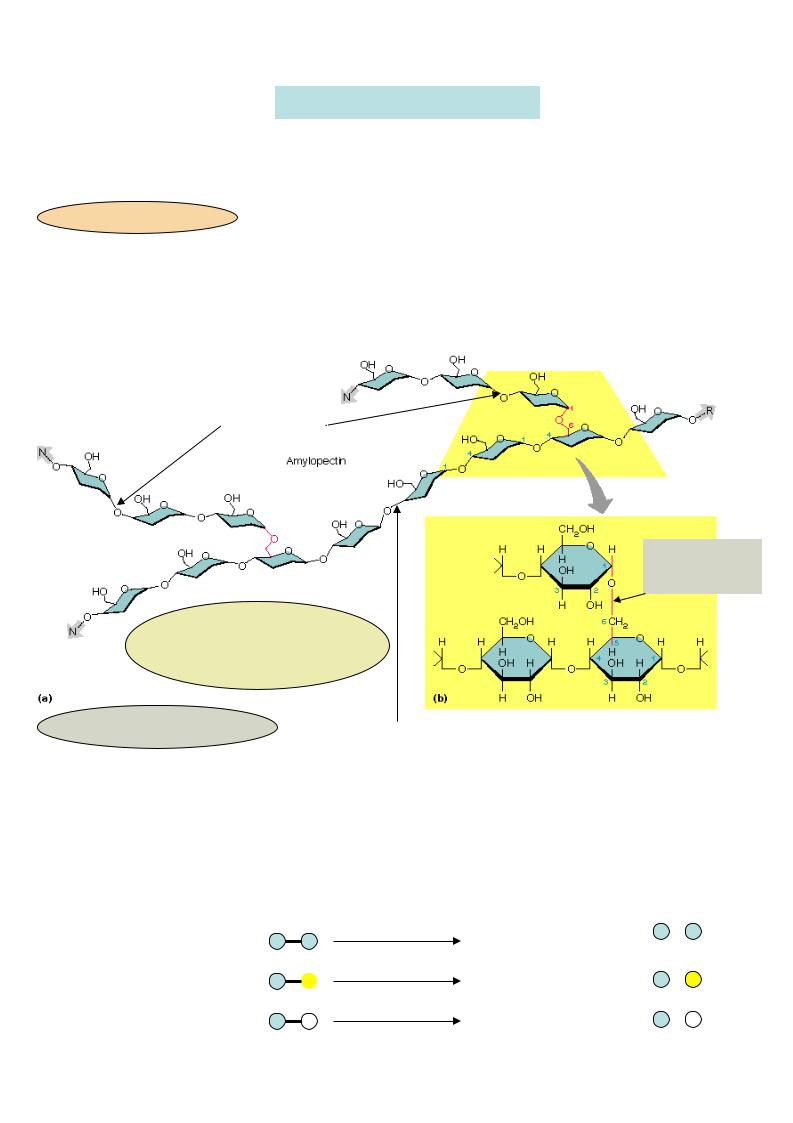
Крахмал – разветвленный гомополисахарид, состоит из мономера глюкозы, Мономеры линейных участков (амилозы)
соединены 1,4-гликозидными
связями, а в местах разветвления
– 1,6-гликозидными связями.
Гликоген – полисахарид, по структуре подобный крахмалу, но более разветвленный (т.е. содержит больше 1,6-гликозидных связей, а значит большее количество концевых мономеров, что облегчает его распад ).
1,6-гликозидная  связь
связь
1,4-гликозидная связь 50
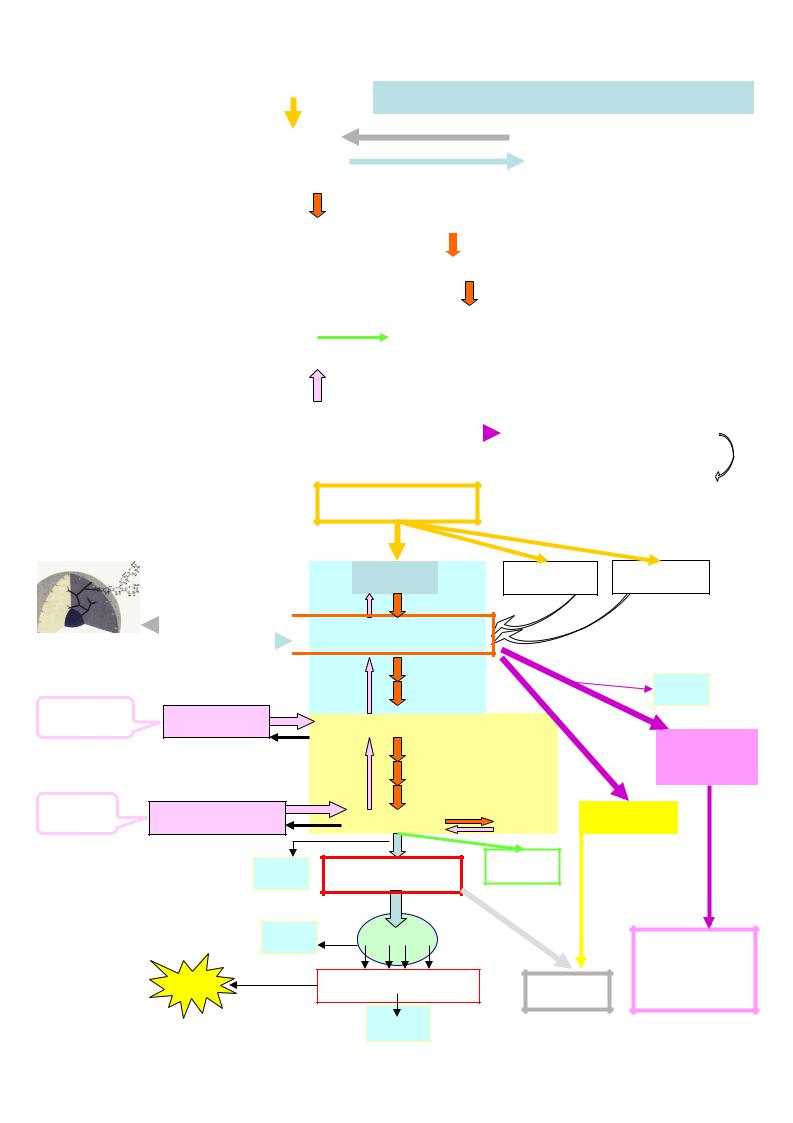
I. Основные процессы в обмене углеводов
1.Переваривание углеводов
2.Синтез гликогена - гликогенез
3.Распад гликогена - гликогенолиз
4. Дихотомический распад глюкозы - непрямое окисление глюкозы при делении ее молекулы на две триозы:
4.1. Анаэробный гликолиз - окисление глюкозы в бескислородных условиях с образованием двух молекул лактата и 2 АТФ 
4.2. Аэробный гликолиз - окисление глюкозы в кислородных условиях с образованием двух молекул пирувата и 8 АТФ
4.3. Спиртовое брожение - превращение глюкозы в этиловый спирт под действием микроорганизмов (дрожжей)
5.Глюконеогенез - синтез глюкозы из неуглеводов: аминокислот (белков), глицерина (липидов), пирувата, лактата
6.Апотомический распад глюкозы - прямое окисление глюкозы – пентозо-фосфатный путь 
7.Взаимопревращения моносахаров – включение других гексоз в обмен глюкозы
Углеводы пищи |
|
|
глюкоза |
фруктоза |
галактоза |
гликоген 
 глюкозо-6-фосфат
глюкозо-6-фосфат
липиды |
|
|
|
СО2 |
глицерин |
3-фосфоглицеральдегид |
рибозо- |
||
|
|
|
|
|
|
|
|
|
5-фосфат |
белки |
аминокислоты |
пируват |
лактат НАДФН+ |
|
|
СО2 |
ацетил-SКоА |
этанол |
|
|
|
|
||
|
СО2 |
ЦТК |
|
нуклеотиды |
|
АТ |
дыхательная цепь |
липиды |
коферменты |
|
Ф |
ДНК, РНК |
||
51 |
|
Н2О |
|
|
II. Переваривание углеводов
Переваривание углеводов – это гидролиз олиго- и полисахаридов, т.е. расщепление гликозидных связей под действием воды с помощью гидролитических ферментов.
Ротовая полость |
|
Начинается переваривание углеводов в ротовой полости, где действует |
α-амилаза |
слюны (рН opt = 6,7), катализирующая гидролиз 1,4-гликозидных связей в молекуле крахмала. Этот фермент является эндоамилазой, т.е. расщепляет связи, удаленные от
конца цепи. В результате образуются |
олигосахариды (декстрины) и небольшое |
|||
количество дисахарида мальтозы. |
|
|||
|
|
|
||
|
α-амилаза |
|
|
|
|
слюны |
|
|
|
|
|
|
|
|
|
|
|
|
|
1,6-
гликозидаза
В желудке (рН 1,5-2) переваривание углеводов
не происходит
|
Тонкий кишечник |
|
|
|
|
|
|
|
|
панкреатическая |
|
|||
|
|
|
||
|
|
α-амилаза |
|
|
В верхнем отделе тонкой кишки действует |
панкреатическая α-амилаза, которая |
|||
гидролизует 1,4-гликозидные связи, последовательно отщепляя молекулы мальтозы от декстринов. 1,6-гликозидные связи в декстринах гидролизуют 1,6-гликозидазы.
На дисахариды в кишечнике действуют соответствующие ферменты ( |
мальтаза, |
сахараза и лактаза), расщепляющие их до моносахаридов: |
|
мальтоза
|
мальтаза |
сахароза |
сахараза |
глюкоза + глюкоза |
+ |
глюкоза + фруктоза |
+ |
лактоза |
лактаза |
глюкоза + галактоза |
+ |
52 |
Превращение глюкозы в тканях
АТФ АДФ
|
гексокиназа |
глюкоза |
глюкокиназа |
глюкоза-6-фосфат |
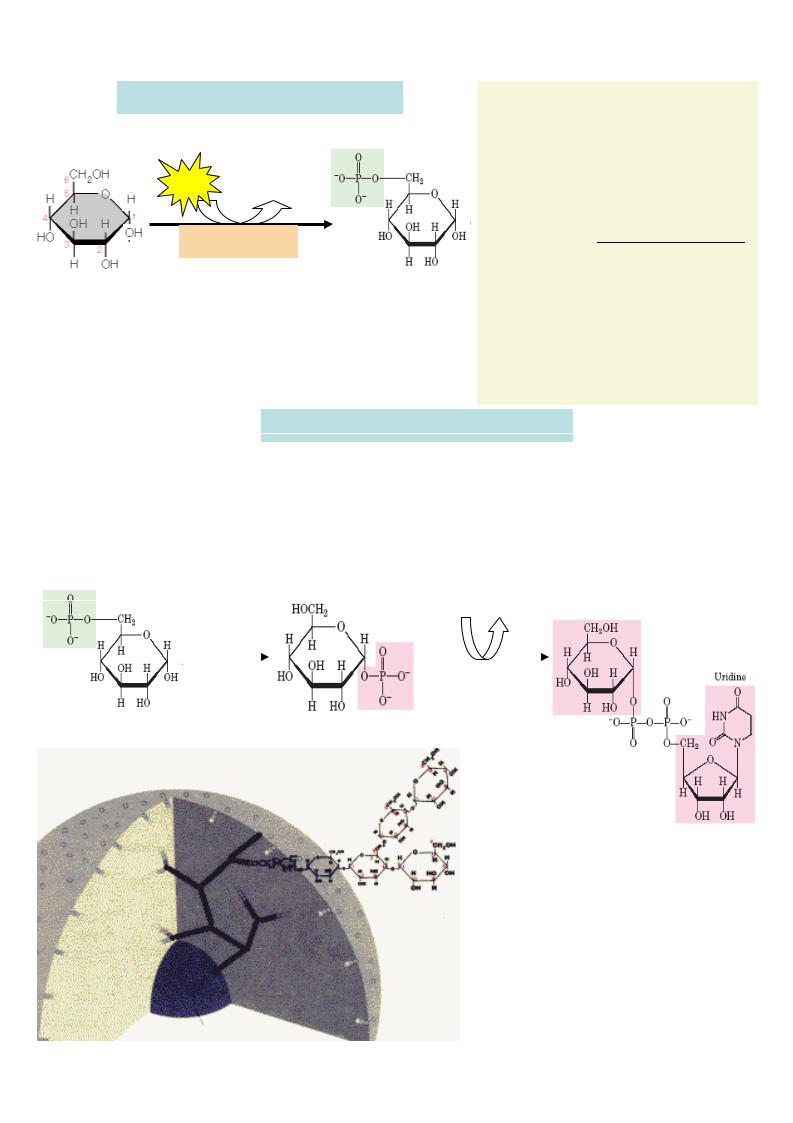
Эта реакция «запирает» глюкозу в клетке, т.к. клеточная мембрана непроницаема для фосфорилированной глюкозы.
Первая реакция превращения глюкозы в тканях – активация
глюкозы путем фосфорилиро-
вания с участием АТФ. Катализирует ее во многих тканях гексокиназа, а в клетках печени – глюкокиназа .
Глюкокиназа специфична для глюкозы и работает при высоких ее концентрациях ( КM
= 12ммоль/л), а гексокиназа
фосфорилирует разные гексозы и ингибируется глюкозо-6- фосфатом (КM = 0,1ммоль/л).
III. Синтез гликогена - гликогенез
Гликоген – животный гомополисахарид, резервная форма глюкозы , в основном депонируется в печени и скелетных мышцах . Гликоген животных, как и амилопектин растений, представляет собой разветвленный полимер глюкозы , в котором остатки глюкозы соединены α(1→4)-гликозидной связью. Связи в точках ветвления находятся в положении α(1→6) примерно каждого 10-го остатка. Разветвленная структура создает большое количество концевых мономеров, что способствует работе ферментов, отщепляющих или присоединяющих глюкозу при его синтезе или распаде.
|
|
УТФ ФФ |
|
|
|
|
|
фосфо- |
УДФ-глюкозо- |
||
глюкомутаза |
пирофосфорилаза |
||
глюкоза-6-фосфат глюкозо-1-фосфат
УДФ-глюкоза
|
|
Хранение в организме собственно |
|
|
глюкозы неприемлемо из-за ее |
|
|
высокой растворимости: высокие |
|
|
концентрации глюкозы создают в |
|
|
клетке высокогипертоническую |
|
|
среду, что приводит к притоку |
|
|
воды. Напротив, нерастворимый |
53 |
Гранулы гликогена |
гликоген осмотически почти |
неактивен и компактен. |
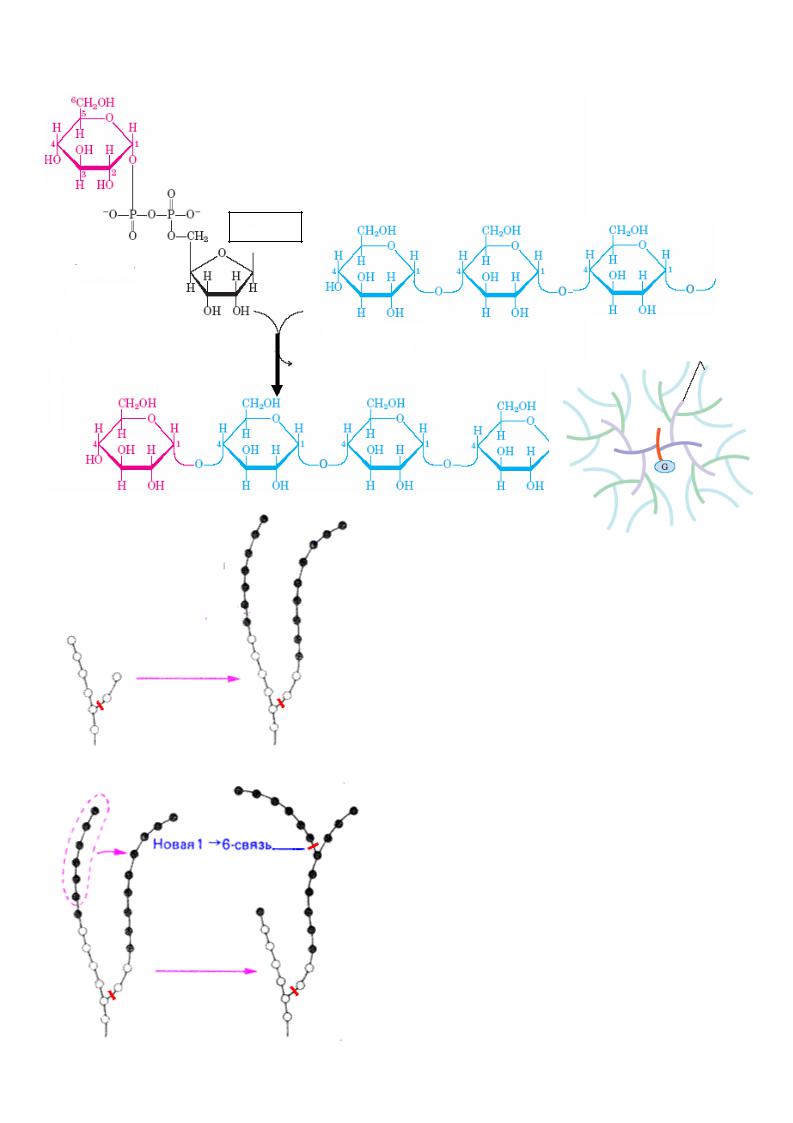
Удлинение цепи линейных полимеров путем образования 1,4-гликозидных связей
урацил
УДФ-глюкоза
олигосахарид - «затравка» гликогена (праймер), гликоген- содержащий n молекул глюкозы (n>4)
синтаза
УДФ
гликоген, содержащий n+1 молекул глюкозы
а) удлинение цепи
гликогенсинтаза
б) ветвление
ветвящий фермент
Удлинение цепи катализируется гликогенсинтазой. Так как образование гликозидных связей между сахарами является
эндоэргической реакцией, вначале в реакции глюкозо-1-фосфата с уридинтрифосфатом (УТФ) образуется активированный предшественник — УДФ-глюкоза. После этого остаток глюкозы легко переносится с УДФ-глюкозы на «затравку» гликогена. Когда растущая цепь достигает определенной длины (>11 остатков), специальный ветвящий фермент гликогена
(1,4→1,6-трансгликозидаза ) катализирует перенос концевого олигосахарида, состоящего из 6-7 остатков, на 6-ОН остаток глюкозы той же или другой цепи гликогена с образованием точки ветвления [α(1→6)-
связи]. Дальнейшее удлинение этого |
|
фрагмента осуществляется |
|
гликогенсинтазой, образующей α(1→4)- |
|
связи. |
54 |
|
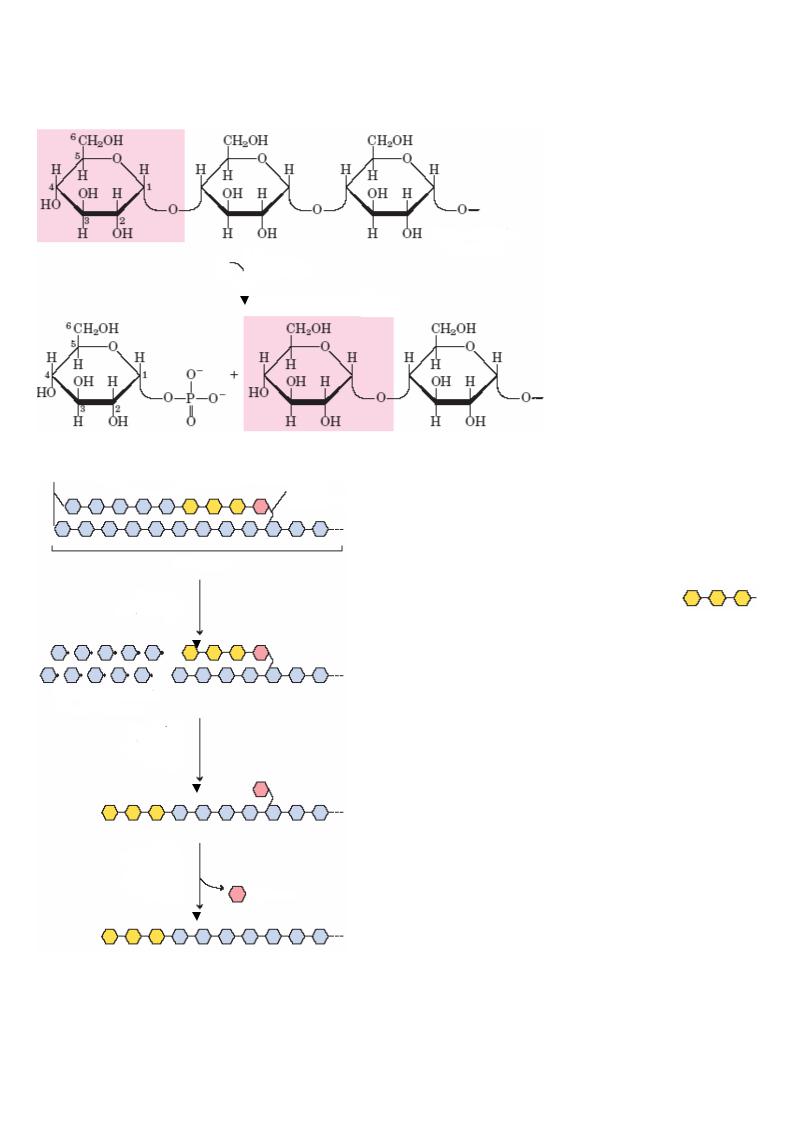
IV. Распад гликогена в тканях |
|
|
Гликоген печени никогда |
||||||||||||
|
|
|
|
|
|
|
|
|
|
|
не расщепляется |
|||||
|
|
|
|
|
|
|
|
|
|
|
||||||
|
|
|
|
|
|
|
|
|
|
|
полностью. Как правило, |
|||||
|
|
|
|
|
|
|
|
|
|
|
укорачиваются |
только |
||||
|
|
|
|
|
|
|
|
|
|
|
невосстанавливающие |
|||||
|
|
|
|
|
|
|
|
|
|
|
концы древовидной |
|||||
|
|
|
|
|
|
|
|
|
|
|
структуры путем |
|||||
|
Н3РО4 |
|
Гликоген ( n молекул глюкозы) |
|
фосфоролиза, т.е. |
|||||||||||
|
|
|
||||||||||||||
гликоген- |
|
|
|
|
|
|
|
присоединения |
|
|
|
|||||
фосфорилаза |
|
|
|
|
|
|
|
фосфорной кислоты по |
||||||||
|
|
|
|
|
|
|
||||||||||
|
|
|
|
|
|
|
|
|
|
|
месту разрыва связи. |
|||||
|
|
|
|
|
|
|
|
|
|
|
|
Гликогенфосфорилаза |
||||
|
|
|
|
|
|
|
|
|
|
|
|
отщепляет от |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
неразветленных цепей |
||||
|
|
|
|
|
|
|
|
|
|
|
|
гликогена по одному |
||||
Глюкозо-1-фосфат |
|
Гликоген ( n -1 молекул глюкозы) |
|
|
мономеру в виде |
|||||||||||
|
|
|
глюкозо-1-фосфата |
|||||||||||||
|
|
|
|
|
|
Гликогенфосфорилаза |
|
|||||||||
|
|
|
|
|
1,6-связь |
катализирует фосфоролиз |
||||||||||
|
|
|
|
|
|
1,4-связей, т.е. расщепление линейных |
|
|
|
|||||||
|
|
|
|
|
|
полимеров. Разрыв |
1,6-связей катализирует |
|||||||||
гликоген |
|
|
деветвящий фермент, который обладает: |
|||||||||||||
|
|
|
гликозилтрансферазной активностью , т.е. |
|||||||||||||
|
|
|
|
|
|
|
|
|
|
|
||||||
гликоген- |
|
|
|
|
переносит олигосахарид из трех глюкоз |
|
|
|
||||||||
фосфорилаза |
|
|
|
|
в конец неразветвленной цепи, при этом остается |
|||||||||||
|
|
|
|
|
|
одна молекула глюкозы, связанная 1,6-связью; |
||||||||||
|
|
|
|
|
|
|
|
1,6-гликозидазной активностью |
|
, т.е. |
||||||
молекулы глюкозо-1-фосфата |
расщепляет 1,6-связи с образованием свободной |
|||||||||||||||
глюкозы и линейного полимера, на который |
||||||||||||||||
деветвящий |
|
|
|
|
||||||||||||
|
|
|
|
далее действует гликогенфосфорилаза, отщепляя |
||||||||||||
|
|
|
||||||||||||||
фермент |
|
|
|
|
||||||||||||
|
|
|
|
по одной молекуле глюкозо-1-фосфата. |
|
|
|
|||||||||
гликозил- |
|
|
|
|
|
|
|
|||||||||
трансферазная |
|
|
|
|
Распад гликогена в печени и в мышцах |
|
|
|
||||||||
активность |
|
|
|
|
отличается только одной реакцией, |
|
|
|
||||||||
|
|
|
|
|
|
|||||||||||
|
|
|
|
|
|
|
|
|
||||||||
|
|
|
|
|
|
обусловленной наличием |
только в печени |
|||||||||
деветвящий |
|
|
|
|
глюкозо-6-фосфатазы, которая «освобождает» |
|||||||||||
|
|
|
|
|||||||||||||
|
|
|
|
глюкозу от фосфата |
(своеобразного якоря, |
|||||||||||
фермент |
|
|
|
|
||||||||||||
1,6-гликозидазная |
|
|
|
глюкоза |
который не выпускает глюкозу из клетки), в |
|||||||||||
активность |
|
|
|
результате чего глюкоза из печени выходит в |
||||||||||||
|
|
|
|
|
||||||||||||
линейный (1,4-) полимер - |
кровь. Таким образом, |
мобилизация гликогена |
||||||||||||||
печени поддерживает постоянную |
|
|
|
|||||||||||||
субстрат для дальнейшего |
концентрацию глюкозы в крови. |
Функция |
||||||||||||||
действия гликогенфосфорилазы |
гликогена мышц – обеспечение самой мышцы |
|||||||||||||||
55 |
|
|
|
|
|
метаболическим топливом. |
|
|
|
|||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|

Регуляция гликогенолиза и гликогенеза (синтеза и распада гликогена)
Синтез и распад гликогена – два противоположных процесса, их одновременное протекание в клетке бессмысленно, поэтому происходит переключение этих метаболических путей путем изменения активности двух ключевых ферментов –
гликогенсинтазы и гликогенфосфорила с помощью |
|
|
инсулин |
||
их фосфорилирования и дефосфорилирования. |
||
|
Синтез гликогена активируется в период пищеварения (1-2 часа после приема углеводной пищи). В это время вырабатывается гормон поджелудочной железы инсулин, который активирует гликогенсинтазу путем ее дефосфорилирования (см. раздел «Гормональная регуляция обмена веществ»). В результате глюкоза, всосавшаяся в кровь из кишечника, депонируется в печени в виде гликогена.
протеинфосфатаза
Н2О
гликоген- |
гликоген- |
фосфорилаза |
фосфорилаза |
неактивная |
активная |
|
АТФ АДФ
протеинкиназа
глюкагон адреналин
гликоген- |
гликоген- |
синтаза |
синтаза |
неактивная |
активная |
|
Н2О
протеинфосфатаза
глюкозо- 6-фосфат инсулин глюкагон
Распад (мобилизация) гликогена
происходит, главным образом, в период между приемами пищи , а также во время физической работы. В постабсорбтивном периоде (при снижении концентрации глюкозы в крови) усиливается выработка другого гормона поджелудочной железы глюкагона, который активирует гликогенфосфорилазу путем фосфорилирования. При этом происходит распад гликогена в печени и глюкоза выходит в кровь. Аналогично в мышцах действует адреналин, который активирует распад гликогена при мышечной работе.
Болезни гликогена
Это наследственные заболевания – энзимопатии, связанные со снижением или отсутствием активности ферментов синтеза или распада гликогена.
Гликогенозы – болезни накопления гликогена, вследствие чего возникает гипогликемия и ее последствия (например, болезнь Гирке – недостаточность глюкозо-6-
фосфатазы).
Агликогенозы – нарушение синтеза гликогена, в результате чего наблюдается |
56 |
снижение его содержания в тканях и также гипогликемия. |

|
|
|
|
глюкоза |
|
|
|
|
|
|
V. Дихотомический распад глюкозы |
|||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
– непрямое окисление глюкозы. |
|||
|
|
|
|
|
|
|
гликолиз |
|
|
|
|
|||||||
|
|
|
|
|
|
|
|
|
|
|
Независимо от условий, |
|
|
|||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
дихотомическое деление |
6-ти |
||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||||
|
|
|
|
2 пируват |
|
|
|
|
|
|
||||||||
анаэробные условия |
|
|
|
|
анаэробные |
углеродной молекулы глюкозы с |
||||||||||||
в дрожжах |
|
|
|
|
|
|
|
|
|
|
условия |
образованием двух 3-х углеродных |
||||||
|
|
|
|
|
аэробные |
|||||||||||||
|
|
|
|
|
|
|
|
|
|
|
||||||||
|
|
|
|
|
|
|
|
|
|
|
молекул пирувата происходит |
|||||||
2 этанол +2СО2 |
|
|
|
|
|
|
условия |
|
|
|
2 лактат |
|
||||||
|
|
|
|
|
|
2СО2 |
|
|
одинаково . Далее, в зависимости |
|||||||||
|
|
|
|
|
|
|
|
|
|
|
от условий, пируват различными |
|||||||
|
|
|
|
|
|
|
|
|
|
|||||||||
|
|
|
2 ацетил-КоА |
|
|
|
|
|
|
путями превращается либо в |
|
|||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
лактат, либо в ацетил-КоА, или в |
|||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||||
|
|
|
|
4СО2 + Н2О |
|
|
|
|
|
|
этиловый спирт (в дрожжах). |
|
||||||
|
|
|
|
|
|
|
|
|
|
Анаэробный гликолиз |
|
|||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|||
Анаэробный гликолиз – это ферментативный распад глюкозы до молочной |
кислоты |
|||||||||||||||||
(лактата) без потребления кислорода (см. схему на стр.). Все реакции гликолиза |
|
|||||||||||||||||
происходят в цитозоле. |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||||
Происходит в мышцах в первые минуты интенсивной работы, |
эритроцитах (нет |
|||||||||||||||||
митохондрий), в различных органах при недостаточном снабжении их кислородом |
|
|||||||||||||||||
(гипоксия), а также в |
опухолевых клетках . В этих условиях гликолиз является |
|
||||||||||||||||
единственным способом получения энергии для синтеза АТФ из АДФ и Фн. |
|
|
||||||||||||||||
1 стадия - энергопотребляющая |
, в которой глюкоза дважды фосфорилируется |
|
||||||||||||||||
(активируется) |
с затратой |
2АТФ |
и дихотомически делится на два |
|
|
|||||||||||||
фосфорилированных С |
3 фрагмента - фосфотриозы: фосфодиоксиацетон и |
3- |
|
|||||||||||||||
фосфоглицериновый альдегид, которые взаимопревращаются, но во вторую стадию |
|
|||||||||||||||||
непосредственно включается 3-фосфоглицериновый альдегид. |
|
|
|
|||||||||||||||
2 стадия - энергогенерирующая стадия |
(гликолитическая оксидоредукция) |
, в |
||||||||||||||||
которой происходит окисление |
(дегидрирование) |
3-фосфоглицеральдегида и |
|
|||||||||||||||
восстановление НАД+ в НАДН. Кроме того, во второй стадии происходят две реакции субстратного фосфорилирования (7-я и 10-я реакции), каждая из них дает 2АТФ в расчете на одну молекулу глюкозы (т.к. в результате дихотомии образовалось две
фосфотриозы и каждая далее превращается в пируват). Таким образом, во второй |
|||||||||||
стадии образуется 4 молекулы АТФ. |
|||||||||||
|
|
|
|
|
|
|
|
|
|
|
НАДН Н +, образовавшийся при окислении 3- |
|
|
глюкоза |
|
|
|
|
|
фосфоглицеринового альдегида (6-я реакция), |
|||
|
|
|
|
|
|
|
|
|
|
|
в анаэробных условиях не может окисляться в |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
дыхательной цепи, поэтому восстанавливает |
|
|
|
|
|
2НАД+ |
|
|
|
|||
|
|
|
|
|
|
|
|
пируват до молочной кислоты (11-я реакция), |
|||
|
|
|
|
|
|
|
|
|
|
|
что обеспечивает регенерацию окисленного |
|
|
|
|
2НАДНН+ |
|
|
|
|
НАД+ в отсутствие кислорода. |
||
2 пируват |
|
|
2 лактат |
|
|||||||
|
|
в отсутствие О2 |
|
Всего превращение глюкозы в лактат |
|||||||
|
|
|
|
|
|
||||||
57 |
|
|
|
|
|
|
|
|
|
|
сопровождается образованием (2х2-2)=2АТФ. |
|
|
|
|
|
|
|
|
|
|
|
|
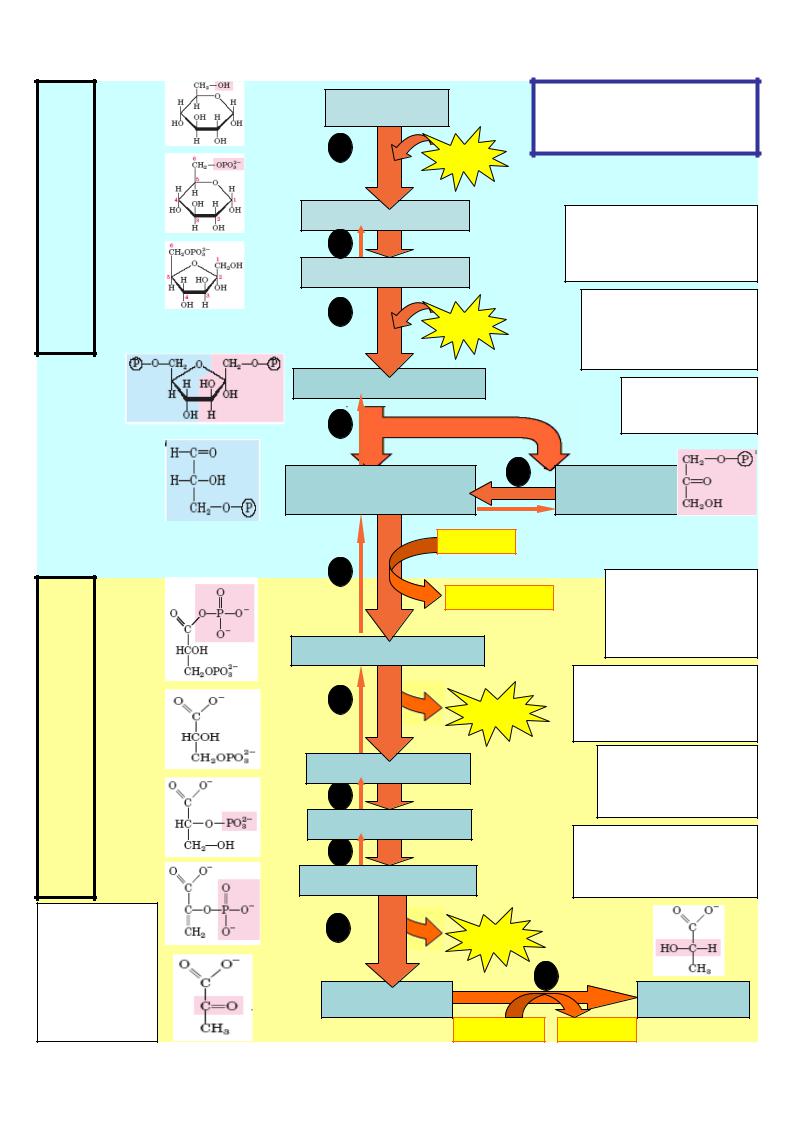
энергопотребляющая стадия
Энергогенерирующая гликолитическая (
Реакция 11
Восстановление пирувата в лактат (используется НАДН Н+ , обоазующийся в 6-й реации)
глюкоза |
|
|
Анаэробный |
1 |
АТ |
|
гликолиз |
|
|
||
гексокиназа |
Ф |
|
|
глюкозо-6-фосфат |
|
Реакции 1-3 |
|
2 |
|
|
Активация глюкозы путем |
|
|
фосфорилирования |
|
фруктозо-6-фосфат |
|
затрачено 2 моля АТФ |
|
3 |
АТ |
|
Реакция 4 |
|
Дихотомия – деление |
||
фосфофрукто- |
Ф |
|
6-углеродной молекулы |
киназа |
|
|
нп 2 фосфотриозы |
фруктозо-1,6-дифосфат |
|
Реакция 5 |
|
|
|
|
|
4 |
|
|
Изомеризация |
|
|
триозофосфатов |
|
3-фосфоглицериновый |
5 |
фосфодиокси- |
|
альдегид |
|
|
ацетон |
|
2 НАД+ |
|
|
6 |
|
|
Реакция 6 |
|
2 НАДН Н+ |
||
|
Образование 2 молей |
||
|
|
|
НАДНН+ и |
|
|
|
макроэргического |
(2)1,3-дифосфоглицерат |
|
соединения |
|
7 |
|
|
Реакция 7 |
2АТ |
|
Субстратное |
|
|
фосфорилирование- |
||
|
Ф |
|
|
|
|
образование 2 молей АТФ |
|
(2)3-фосфоглицерат |
|
Реакции 8 и 9 |
|
|
Образование |
||
8 |
|
|
макроэргического |
|
|
соединения (и воды) |
|
(2)2-фосфоглицерат |
|
||
|
Реакция 10 |
||
9 |
|
|
|
|
|
Субстратное |
|
(2)фосфоенолпируват |
|
фосфорилирование - |
|
|
образование 2 молей АТФ |
||
|
|
|
|
10 |
2АТ |
|
|
пируваткиназа |
Ф |
|
|
|
|
|
|
(2) пируват |
11 |
(2)лактат |
|
||
2 НАДН |
2 НАД+ |
58 |
|

1 - гексокиназа
2 - глюкозо-6-фосфат изомераза
3 – фосфофруктокиназа
4 – альдолаза
5 – триозофосфатизомераза
6 – глицеральдегидфосфатдегидрогеназа
7 – фосфоглицераткиназа
8 – фосфоглицеромутаза
9 – енолаза
10пируваткиназа
11лактатдегидрогеназа
стадия 1
стадия 2
Из 11 реакций анаэробного гликолиза три реакции необратимы, остальные могут протекать как в прямом, так и в обратном направлениях. Необратимыми являются: 1 – гексокиназная; 3 – фосфофруктокиназная;
10 – пируваткиназная.
Ферменты, катализирующие необратимые реакции, являются регуляторными, от их активности зависит скорость всего метаболического процесса.
|
|
|
|
|
|
|
|
|
|
|
VI. Аэробный гликолиз и |
|
||||
|
|
|
|
|
|
|
|
|
|
|
|
глюконеогенез |
|
|||
|
|
|
|
|
|
глюкоза |
|
|
||||||||
|
|
|
|
|
|
|
|
|
|
Аэробный гликолиз – это окисление глюкозы |
||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
до двух молекул пирувата в кислородных |
|
|
|
|
|
|
|
|
|
|
|
2НАД+ |
|
|
|
условиях. При этом образуется |
2 моля |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
гликолитического НАДН Н + |
, которые в |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
аэробных условиях восстанавливают |
|
|
|
|
|
|
|
|
|
2НАДНН+ |
|
|
ЦПЭ |
|||||
|
|
|
|
|
|
|
|
|
|
кислород в цепи переноса электронов (ЦПЭ) |
||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|||
|
|
2 пируват |
|
|
|
|
|
|||||||||
|
|
|
|
|
|
|
|
|
|
с образованием воды. Одновременно в |
||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
результате окислительного |
|
в присутствии |
|
|
|
|
|
|
|
|
|
|
|
|
||||
кослорода |
|
|
|
|
|
|
|
|
|
|
|
фосфорилирования образуется 2х3=6 молей |
||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
АТФ. Учитывая субстратное |
|
|
|
ацетил-КоА |
|
|
|
|
||||||||||
|
|
|
|
|
фосфорилирование гликолиза |
(2АТФ), |
||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
выигрыш энергии при превращении глюкозы |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
в пируват составляет 8 АТФ (см. схему на |
|
|
|
СО2 |
|
|
Н2О |
|
|
|
|
|
|
|||||
|
|
|
|
|
|
|
|
стр. 55). |
|
|||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Далее пируват в присутствии кислорода окисляется в ацетил-КоА с образованием 3АТФ (за счет окислительного фосфорилирования), а ацетил-КоА окисляется в ЦТК с образованием СО 2, Н 2О и 12АТФ. Таким образом, при окислении пирувата до углекислого газа и воды образуется 15АТФ, а т.к. из глюкозы образуется 2 моля пирувата, то они при «сгорании» дают 30АТФ. Всего полное окисление глюкозы в кислородных условиях дает 30+8=38 молей АТФ.
Аэробное окисление глюкозы является намного более выгодным процессом, т.е. глюкоза расходуется более экономно (медленно) по сравнению с анаэробным гликолизом.
Поэтому при переключении с анаэробного на аэробный путь снижается скорость расходования глюкозы и прекращается накопление лактата ( эффект Пастера ). Молекулярный механизм этого эффекта состоит в конкуренции между дыханием и
гликолизом за АДФ, необходимым для образования АТФ.
59