II. Репарация
Передача генетической информации в неискаженном виде - важнейшее условие сохранения вида. Это обеспечивается за счет комплементарности. Однако, возможны как ошибки репликации, так и неблагоприятное действие окружающей среды (радиация, УФ-облучение, химические агенты).
Cуществует специальная система мониторинга точности спаривания нуклеотидов, причем осуществляется двойная проверка:
-при включении ДРН в растущую цепь;
-после включения ДРН путем удаления ошибочных нуклеотидов (ошибки происходят
не чаще, чем 1 раз на 10 пар оснований).
Причины спонтанных нарушений
ошибки репликации
депуринизация вследствие непрочности N-гликозидной связи в пуринах дезаминирование (Ц У, А Г, Г Х)
Причины индуцированных нарушений
алкилирование азотистых оснований (алкилирующие агенты применяются как ингибиторы новообразований)
образование сшивок - пиримидиновых димеров между соседними основаниями (тимином) под действием УФ-облучения
сахаро-фосфатный
ДНК |
остов |
пары оснований, |
|
|
связанные |
водородными
связями
повреждение
удаление поврежденного участка
ДНК-полимераза I строит новую копию
ДНК-лигаза сшивает разрыв
Репарация – удаление поврежденных участков ДНК или ошибочно встроенных нуклеотидов. Ключевой частью всех репаративных систем является распознавание дефекта в структуре ДНК, который или сразу восстанавливается, или маркируется специальными белками.
Этапы репарации:
1) узнавание места повреждения и раскручивание участка ДНК в области повреждения 2) однонитевой разрыв ДНК с 5 –конца на
расстоянии около 20 п.о. от места повреждения
УФ-специфической эндонуклеазой
3) достройка (“латка”) поврежденной цепи с помощью ДНК-полимеразы 1
4)отщепление поврежденного участка за счет экзонуклеазной активности ДНК-полимеразы I
5)сшивание 3-конца латки с 5-концом
основной цепи ДНК-лигазой
В клетке действует несколько систем репарации, позволяющих сохранить генетическую информацию. Нарушение работы таких систем приводит к заболеваниям, например, к пигментной ксеродермии – аутосомно-рецессивному наследственному заболеванию. Существует две основные клинические формы проявления этого заболевания: одна сопровождается дегенеративными изменениями в глазах и коже, а другая, кроме того, характеризуется неврологической дегенерацией. Тяжелым осложнением этого заболевания является рак кожи. Причины развития пигментной ксеродермии связывают с дефектами ферментов репарации
тиминовых димеров |
(в частности, нарушение синтеза |
УФ-специфичной |
эндонуклеазы). |
|
|
|
III. Генные мутации |
|
Изменения генетической программы ДНК клеток называются мутациями. Различают хромосомные мутации (изменение числа хромосом, хромосомные абберации) и молекулярные или генные мутации.
Генные мутации - это наследуемые изменения первичной структуры ДНК, которые ведут либо к прекращению синтеза белка, либо к синтезу измененного, дефектного белка.
Мутации в регуляторных участках оперона ведут к нарушению регуляции или прекращению синтеза ДНК.
Существуют следующие варианты генных мутаций: Транзиция - замена пар оснований
Миссенс-мутация - приводит к изменению смысла кодона при замене нуклеотида, а, значит, к синтезу измененного белка. Например, серповидноклеточная анемия: кодон, отвечающий за включение глу в цепь гемоглобина, превращается в кодон вал.
Замена одного нуклеотида не всегда ведет к изменению смысла кодона (т.к. код вырожденный) - такое изменение ДНК фенотипически не проявляется.
Нонсенс-мутация - в результате замены образуется один из терминирующих кодонов, при этом синтез белка прекращается и образуется незавершенный белок.
Делеция - выпадение одной пары или групп оснований
Вставка одной пары или групп оснований. Эти мутации могут быть как с изменением рамки считывания (выпадение или вставка одного или двух нуклеотидов), что ведет к синтезу бессмысленного белка, так и без изменения рамки считывания (три нуклеотида) - синтезируется белок, укороченный на 1 аминокислоту.
Изменение местоположения отдельных участков ДНК.
IV. Молекулярные основы транскрипции
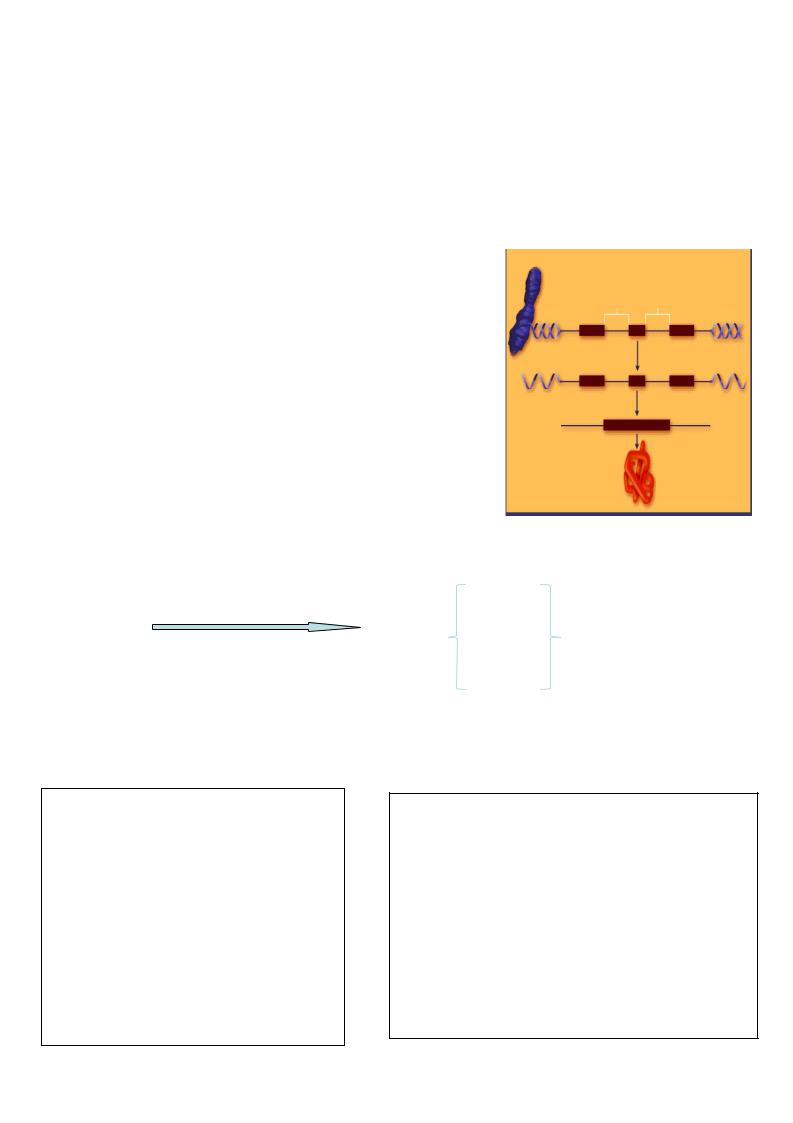
Транскрипция - это синтез всех видов РНК по матрице ДНК, осуществляемый ферментом ДНК-зависимой РНК-полимеразой.
Молекула ДНК, хранящая генетическую информацию, непосредственного участия в синтезе белка не принимает. Возможно лишь снятие с нее копий, которые отправляются в цитоплазму, доставляя генетическую информацию к рибосомам – месту биосинтеза белка.
Каждая такая копия |
– послание ( message, англ.) – |
представляет собой |
м-РНК. Таким образом, |
информация, записанная в последовательности нуклеотидов ДНК копируются (транскрибируются) в виде м-РНК с последующей трансляцией в полипептидную цепь белка. Транскрипция – это первый этап в экспрессии генов.
Экспрессия генов:
транскрипция трансляция
ДНК&
РНК&
белок&
|
Суммарное уравнение транскрипции: |
kАТФ |
ДНК-матрица |
АМФk |
l ГТФ |
РНК-полимеразы |
РНК ГМФl + (k+l+n+m) ФФ |
n ЦТФ |
ЦМФn |
mУТФ |
|
УМФm |
Осуществляется транскрипция с помощью |
ДНК-зависимых РНК-полимераз , для |
работы которых необходимы: матрица – двуцепочечная ДНК, субстраты - АТФ, ГТФ, УТФ, ЦТФ (они же используются как источники энергии), ионы магния.
Упрокариот РНК-полимераза
состоит из |
4 субъединиц, |
образующих |
кор-фермент |
(“сердцевина”), к которому |
присоединяется дополнительный |
белковый |
σ-фактор, |
способствующий более прочному связыванию фермента со специфической промоторной последовательностью на матрице ДНК.
Эукариотические клетки обычно содержат 3 типа РНК-полимераз:
РНК-полимераза I катализирует синтез рибосомальной (рРНК);
РНК-полимераза II катализирует синтез пре-мРНК - ингибируется токсином бледной поганки α-аманитином;
РНК-полимераза III катализирует синтез трпнспортной (тРНК). 132
РНК синтезируется комплементарно и антипараллельно транскрибируемой цепи ДНК. Рост цепи РНК идет только в направлении 5' 3'. Для начала синтеза РНК фермент не нуждается в полиили олигонуклеотидной затравке. Цепь ДНК, на которой идет транскрипция, называется кодирующей, другая - некодирующей.
Принципы транскрипции: |
Отличия ферментов транскрипции и репликации |
|
комплементарность. |
РНК-полимераза не нуждается в праймере |
|
антипараллельность. |
РНК-полимераза сама расплетает материнскую цепь |
|
униполярность. |
РНК-полимераза не обладает нуклеазной активностью |
|
беззатравочность. |
РНК-полимераза перемещается по матрице значительно |
|
асимметричность. |
медленнее, чем ДНК-полимераза |
|
|
|
РНК-полимераз в клетке значительно больше, чем |
|
|
ДНК-полимераз |
|
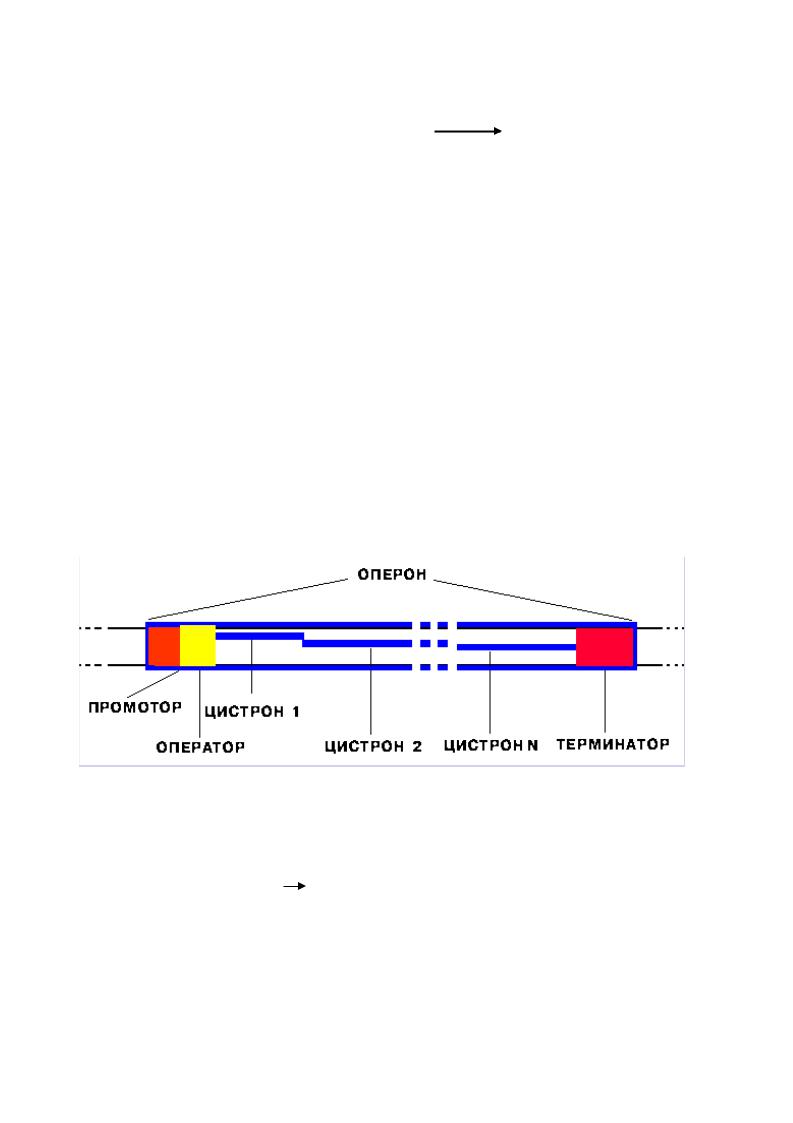
Синтез РНК начинается в особых участках ДНК, называемых |
промоторами, а |
заканчиваются в терминаторах. Участок ДНК, ограниченный промотором и терминатором, называется транскриптон (эукариоты) или оперон (прокариоты). Он включает гены или структурные цистроны, которые несут информацию о структуре полипептидных цепей или нематричных РНК. Транскриптон может содержать один структурный ген или несколько.
Структура оперона
Промотор - участок оперона, служащий для узнавания ферментом РНК-полимеразой. Последовательность оснований по ходу цепи ДНК ниже сайта промотора с направлением 3’ 5’ используется в качестве матрицы для синтеза РНК.
Промоторы прокариот –представляют собой последовательность, состоящую из ~40 нуклеотидов. Узнавание и прочное связывание с РНК-полимеразой происходит на разных участках ДНК. Эти участки отличаются и по первичной, и по вторичной структуре. Путем секвенирования выявили структуру многих промоторов. У
большинства из них имеется общее свойство.
133
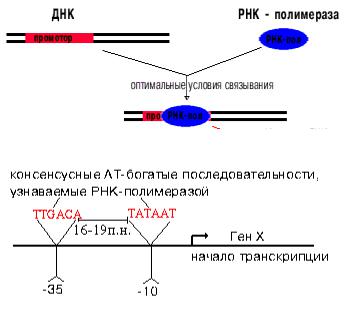
РНК-полимераза узнает промотор,
|
|
|
покрывая |
40-60 пар нуклеотидов. В |
промоторе узнается взаимное |
расположение двух |
AT-богатых |
участков. В каждом из них |
4-6 пар |
нуклеотидов. Центры этих участков |
находятся в |
положениях "-10" и "-35". |
Принципиально важным является расстояние между этими участками. Оно колеблется от 16 до 19 п.н. Искусственное увеличение этого расстояния до 20 п.н. или уменьшение его до 15 п.н. приводит к тому, что РНКполимераза не узнает испорченный промотор.
Для связывания G-фактора РНК-полимеразы с ДНК (сайт инициации) служит « - 35 последовательность» из 6 нуклеотидов, а расплетение двух цепей ДНК происходит в «Pribnowbox», т.е. – «- 10 последовательности». В этих участках много пар А::::Т, между которыми только две водородные связи, поэтому цепи ДНК легко расходятся («плавятся»).
Промоторы эукариот содержат участки, которые не только указывают место начала транскрипции, но и базовую частоту этого процесса.
Большинство генов эукариот содержит в своем промоторном участке специфические последовательности матричной ДНК, которые усиливают (энхансеры) или ослабляют (сайленсеры) уровень экспрессии соответствующих структурных генов, влияя на активность транскрипции, т.е. на количество синтезируемых молекул мРНК и на скорость их образования.
Энхансеры могут повышать эффективность транскрипции генов, на которые они влияют, в десятки и сотни раз. Установлены нуклеотидные последовательности энхансеров для ряда ферментов (алкогольдегидрогеназы, химотрипсина), гормонов (инсулина), иммуноглобулинов.
Для транскрипции требуется не только РНК-полимераза, но и другие белки – основные факторы транскрипции. У эукариот таким фактором является ТАТА-связывающий белок, который взаимодействует с ТАТА-последовательностью (ТАТА-боксом). В этот
|
|
|
|
комплекс встраивается РНК-полимераза и другие |
основные факторы транскрипции. |
Главная задача такого комплекса – «распаковать» |
нуклеосому, а кроме того создать |
условия сорбции |
специфических факторов транскрипции |
, которые могут |
активироваться после взаимодействия с определенными веществами, напрмер, гормонами.
Цистрон - последовательность нуклеотидов ДНК, кодирующая один полипептид (в большинстве случаев - белок) или одну tРНК, или одну rРНК.
В большинстве случаев цистроны объединяются в оперон по следующему принципу: закодированные в них белки принимают участие в одной биохимической цепи реакций.
Гены содержат интроны - неинформативные участки и |
экзоны - |
информативные участки. |
|
Терминатор - последний участок транскриптона, служит сигналом об окончании транскрипции.
Первичный транскрипт - РНК, образующаяся при транскрипции - комплементарная копия транскриптона от промотора до терминатора.
|
ген |
|
промотор |
терминатор |
ЭТАПЫ |
ДНК |
|
|
ТРАНСКРИПЦИИ |
|
|
|
|
|
1. Инициация транскрипции
|
кодирующая |
|
|
2. Элонгация |
|
цепь ДНК |
|
транскрипции |
|
|
|
|
|
3. Терминация транскрипции
(первичный транскрипт)
ЭТАПЫ ТРАНСКРИПЦИИ
На всех этапах транскрипции необходимо участие специальных белковых факторов:
А, В – факторов инициации, Е, Н, F – факторов элонгации, ρ-фактор – фактор терминации.
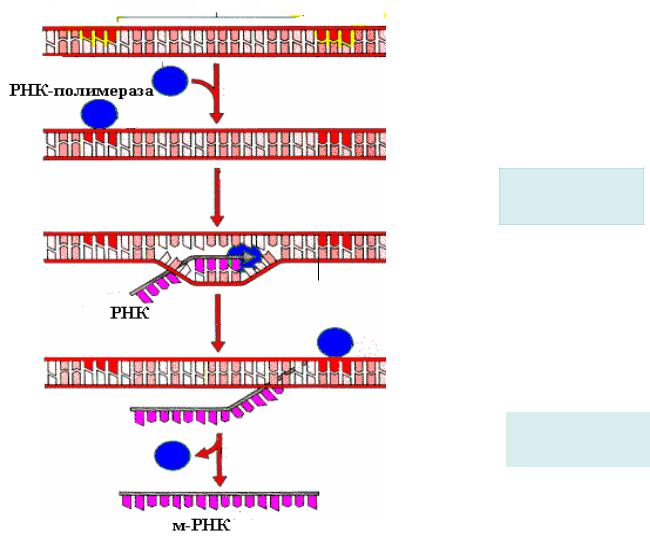
1. Инициация
ДНК-зависимая РНК-полимераза связывается с ДНК и «ищет» промотор, перемещаясь от 3’-конца кодирующей цепи ДНК.
РНК-полимераза находит промотори связывается с ним с помощью σ-фактора. При этом цепь ДНК |
не раскручена. |
|
Цепь ДНК раскручивается с помощью РНК-полимеразы от « |
-10» до « -1» участка, образуя |
открытый промоторный комплекс, который затем с помощью ионов магния еще более расширяется.
Первый нуклеозидтрифосфат, который «ставит» РНК-полимераза, всегда пуриновый: АТФ или ГТФ. Таким образрм, синтез РНК всегда начинается с пуринового нуклеозидтрифосфата (Пур-5-
ФФФ).
На этом этапе действуют ингибиторы - антибактериальные антибиотики
рифамицин и рифампицин, а также противоопухолевый антибиотик
актиномицин D
По мере продвижения кор-фермента происходит расплетание ДНК – как бы движется «пузырь транскрипции», содержащий около 18 п.о. в расплетенном виде. Как только одна п.о. становится раскрученной перед 3 ’-концом растущей цепи РНК, одна п.о. скручивается за «хвостом» РНК-полимеразы.
В результате элонгации образуется гибрид ДНК-РНК.
На этом этапе действует ингибитор элонгации -
стрептолидигин
3. Терминация |
|
|
|
|
|
|
|
|
У E.coli . большинство генов терминируются 2 путями: |
|
1) |
путем формирования структур типа «шпильки» в РНК, в результате чего |
|
ослабляется связь между РНК-полимеразой, матрицей (ДНК) и самой РНК. Эти |
|
структуры образуются благодаря копированию сигналов терминации, которые |
|
представляет собой определенную последовательность в рамках кодирующей |
|
цепи ДНК. Специфичными для терминирующих участков являются |
|
палиндромы |
(обратные повторы) и полиА-последовательности. Транскрипт |
|
с этого участка имеет форму шпильки с концевой |
УУУУ- |
|
последовательностью. |
|
|
2) |
путем узнавания терминирующей последовательности специальным белком- |
|
p-фактором. |
|
|
|
|
|
|
При взаимодействии |
ρ-фактора с |
|
|
|
|
|
|
|
РНК-полимеразой она теряет |
ДНК- |
|
|
|
|
|
|
|
|
|
|
|
|
сродство к ДНК и комплекс |
матрица |
|
|
|
|
|
|
|
|
|
|
|
|
распадается: |
фермент |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
отделяется от ДНК, |
|
|
|
|
|
|
|
|
|
|
|
|
|
терминирующий |
|
|
|
|
|
|
высвобождаются |
фактор и и |
|
|
|
участок |
ДНК |
|
|
|
|
|
|
|
|
|
первичный транскрипт, т.е. РНК |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
После терминации кор-фермент, |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
отделившись от ДНК-матрицы и |
|
|
ρ-фактор |
|
|
|
связавшись с новой молекулой |
первичный транскрипт |
|
|
сигма-фактора, приступает к |
|
|
|
|
|
|
синтезу новой молекулы РНК. |
|
(РНК) |
|
РНК- |
|
полимераза |
|
|
|
|
|
|
|
|
|
|
|
|
137 |
Модификация РНК
У прокариот м-РНК синтезируется сразу в зрелом виде.
У эукариот все первичные транскрипты , образующиеся с помощью РНКполимеразы II, подвергаются процессингу (созреванию) в период между их синтезом и началом реализации соответствующей функции - в роли мРНК, тРНК или рРНК.
|
|
|
|
|
|
Процессинг - сложный процесс, включающий “кэпирование”, |
7-метил- |
реакции расщепления и лигирования |
(сплайсинг), |
присоединение дополнительных концевых нуклеотидов и |
гуанозинтрифосфат |
нуклеозидную модификацию |
|
|
«кэп» |
“Кэпирование” |
- присоединение к 5-концу мРНК |
5’-конец |
|
метилгуанозинтрифосфата (связь 5-5), происходит еще в ядре, в |
|
процессе элонгации, когда длина цепи РНК достигает около 30 |
|
нуклеотидов. РНК-полимераза |
II |
сразу после инициации |
|
«метит» 5’-конец особой меткой, которая получила название |
|
«кэп» (сар – кепка, англ.). |
Она отщепляется от 5’-концевого |
м-РНК |
НТФ последний фосфат, заменяя его остатком ГМФ, |
|
прикрепленной необычной 5’-5’ фосфо-эфирной связью. Далее |
|
метилируется гуанин в положении 7 пуринового кольца. Сар (7- |
|
метилгуанозинтрифосфат) защищает |
м-РНК от действия |
|
экзонуклеаз и отвечает за будущее связывание ее с рибосомой. |
|
Полиаденилирование |
- |
присоединение к 3 ’- концу |
|
синтезированного транскрипта от 20 до 200 остатков АМФ с |
|
|
помощью фермента полиаденилатполимеразы. |
поли-А- |
|
последовательность |
|
Синтезированный «поли-А-хвост» также защищает м-РНК от |
|
3’-конец |
|
экзонуклеаз (с 3’-конца) и способствует выходу м-РНК из ядра. |
|
|
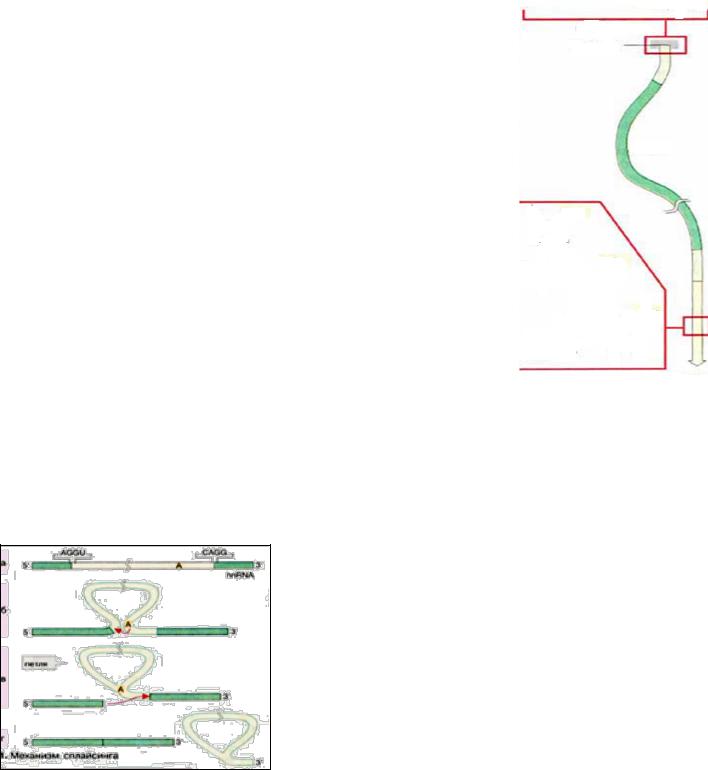
Сплайсинг – точнее вырезание неинформативных участков ( интронов) и сшивание несущих смысловую нагрузку экзонов. При этом экзоны соединяются в том порядке, в каком они присутствовали в первичном транскрипте. Альтернативный сплайссинг, при котором изменяется распределение экзонов одного транскрипта, позволяет увеличить генетическое разнообразие генома (увеличение количества различных белков) без увеличения количества генов.
Все три класса РНК транскрибируются с генов, которые содержат интроны.(до нескольких тысяч нуклеотидов).
Из 25 % длины ДНК, которые кодируют ~ 30 тысяч белков, только 1,1 % - энзоны, а ~24% -интроны. Считается, что присутствие интронов защищает функционально активную часть генома клетки от повреждающего действия химических или физических (лучевых) факторов. 138
V. Трансляция - биосинтез белка
Трансляция- это перевод генетического текста мРНК в линейную последовательность аминокислот в полипептидной цепи белка.
Чтобы аминокислота «узнала» свое место в будущей полипептидной цепи, она должна связаться с транспортной РНК (тРНК), выполняющей адапторную функцию: с одной стороны, «узнает» кодон на мРНК, с другой – связывается с соответствующей этому кодону аминокислотой.
Все т-РНК имеют одну полинуклеотидную цепь, отдельные участки которой образуют спиралевидные петли - “шпильки” за счет водородных связей А-Т и Г – Ц ( 70% п/п цепи), в результате образуется пространственная конфигурация в виде “ клеверного листа”.
Благодаря антикодону -триплету в средней части молекулы тРНК, комплементарному определенному кодону на мРНК, тРНК “читает” текст на м-РНК, а с помощью акцепторного участка на 3-конце - ЦЦА ( обязательного для всех т-РНК) - присоединяет соответствующую аминокислоту с образованием аминоацил-тРНК.
Специфическое взаимное узнавание аминокислоты и тРНК происходит с помощью фермента - аминоацил-тРНК-синтетазы (АРС-азы). Этот фермент узнает и связывает тРНК, а затем переносит аминокислотный остаток на 3’-ОН группу концевого аденозина, присоединяя ее сложно - эфирной связью:
аминокислота + тРНК + ATP |
|
аминоацил-тРНК + АМР + РР |
|
аа-тРНК-синтетаза |
139 |