

Полезные материалы за все 6 курсов / Ответы к занятиям, экзаменам / Биохимия в рисунках и схемах
.pdf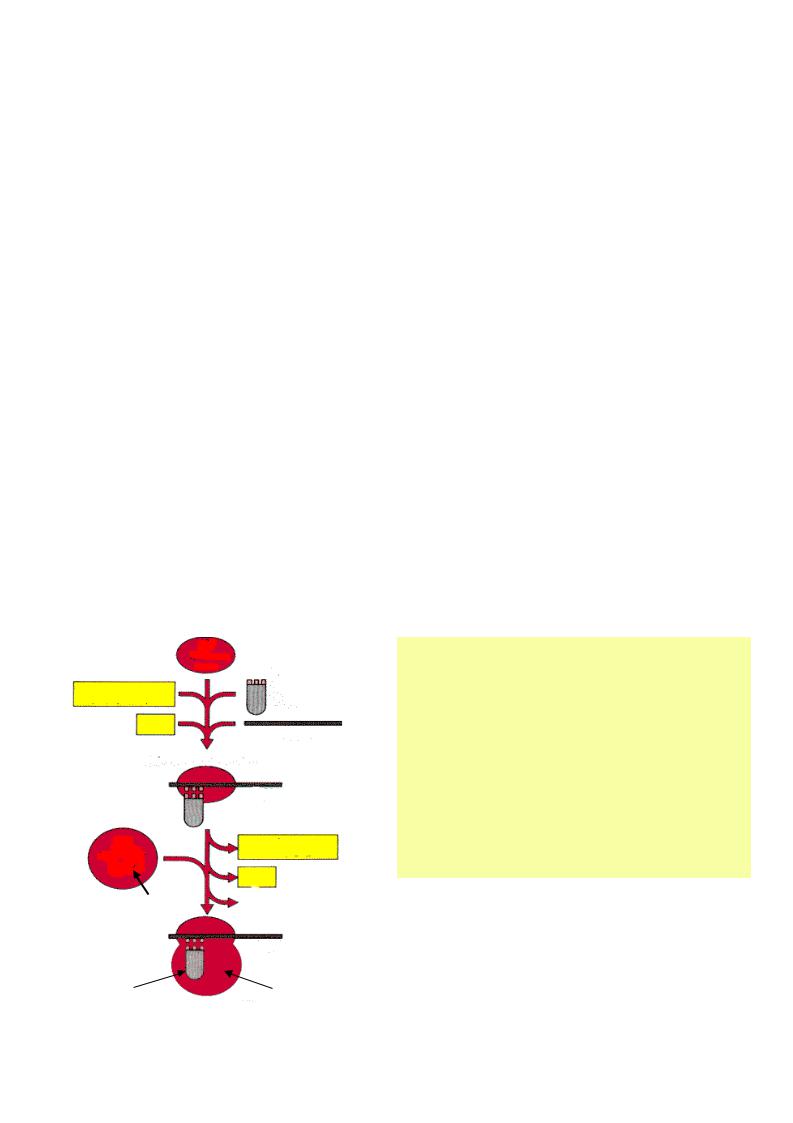
1. ИНИЦИАЦИЯ ТРАНСЛЯЦИИ
Наиболее ответственным моментом трансляции являются точное узнавание самого первого кодона, который соответствует N-концевой аминокислоте будущего белка. От этого зависит правильность установки рамки считывания. При этом оба конца мРНК имеют нетранслируемые участки.
Стартовая точка – инициирующий кодон АУГ, кодирующей метионин, т.е. каждый белок начинается с метионина. Чтобы отличить этот «начальный» метионин от других молекул метионина, которые встраиваются в белки, у клетки есть две разные по структуре тРНК с одинаковыми антикодонами (УАЦ) и только одну из них распознают IF2 (фактор инициации) и только в комплексе с ним эту met-мРНК может принять П-сайт рибосомы. (IF2 обладает ГТФ-азной активностью).
Шаги инициации
• мРНК фиксируется на малой субъединице рибосомы вблизи кэпа
• мет-тРНК вместе с IF2 связывается с малой субединицей рибосомы в П-сайте с участием КЭП-связывающих белков и других факторов инициации.
•у прокариот малая рибосома сразу взаимодействует с участком мРНК, содержащего АУГ или ГУГ (инициирующим), которые соответствуют N- формил мет-т-РНК.
• Малая субъединица, объединенная с мет-т-РНК продвигается по м-РНК к ее 3’-концу с помощью АТФ-зависимого механизма до встречи с кодоном АУГ. По принципу комплиментарности антикодон мет-тРНК (УАЦ) связывается с кодоном АУГ (при этом расщепляется ГТФ), что служит сигналом к объединению малой субъединицы с большой (одновременно в цитоплазму вытесняются белковые факторы). Образуется комплекс: м-РНК-рибосома-метионил-т-РНК, готовый к элонгации.
40S |
|
Факторы инициации у прокариот: |
||
IF1 IF2 IF3 |
|
IF1 ; |
|
|
|
IF2 – обладает ГТФ-азной активностью; |
|||
ГТФ |
метионил-т-РНК IF3 – имеет максимальное сродство к |
|||
мРНК |
малой субъединице. |
|
||
|
|
|||
|
У эукариот факторы инициации eIF1, eIF2… |
|||
|
|
|||
комплекс |
мРНК |
( всего описано до десяти факторов). У эукариот |
||
ингибировать фактор инициации |
eIF -2 могут |
|||
инициации |
||||
|
интерфероны – белки, которые синтезируются |
|||
|
|
|||
|
|
в лимфоидной и других тканях, обладают |
||
IF1 |
IF2 IF3 |
антивирусной и противоопухолевой |
||
60S |
|
активностью. |
|
|
ГДФ |
На рибосоме имеются два участка: |
П-сайт или |
||
cтимулирует |
|
|||
|
пептидильный сайт, служащий для связывания с |
|||
ГТФ-азную активность |
|
|||
фактора IF2 |
мРНК |
удлиняющейся полипептидной цепью; А-сайт |
||
|
|
или аминоацильный, служащий для |
|
|
П-сайт |
А-сайт |
присоединения новой аа-тРНК. На стадии |
||
инициации к П-сайту присоединяется метионил- |
||||
рибосома, |
|
|||
готовая к трансляции |
тРНК, А-сайт – свободен. |
141 |
||


Регуляция&синтеза&белка&!
Концентрация многих белков в клетке непостоянна и изменяется в зависимости от состояния клетки и внешних условий. Это происходит в результате регуляции скоростей синтеза и распада белков. Регуляция экспрессии генов может осуществляться на любой стадии на пути от гена до образования функциональноактивного белка.
Регуляция транскрипции
Транскрипция генов может подавляться или активироваться, следовательно, синтез белков может репрессироваться или индуцироваться. Оперон - это участок ДНК,
кодирующий строение, как правило, функционально связанных белков и содержащий регуляторную зону, контролирующую синтез этих белков. На рисунке приведена схема лактозного оперона:
|
промотор |
оператор |
|
структурные гены |
|
|||||
ген-регулятор |
|
|
|
|
|
|
|
|
||
РНК-полимераза |
|
|
|
|
|
|
Z |
Y |
А |
ДНК |
|
|
|
|
|
|
|||||
ГР |
П |
|
|
О |
||||||
|
|
|
|
|
|
|
транскрипция заблокирована |
|
||
|
|
|
|
|
|
|
|
|||
м-РНК |
белок-репрессор |
|
|
|
|
|||||
|
|
|
|
|
||||||
|
|
|
|
|
|
|
|
|
РНК-полимераза |
|
|
|
|
|
|
|
|
|
|
||
|
|
|
|
|
ИНДУКТОР (лактоза) |
|
|
|||
|
|
|
|
|
|
|
Z |
Y |
А |
ДНК |
|
|
|
|
|
|
|
|
|||
ГР |
П |
|
|
О |
|
|
транскрипция разрешена |
|||
|
|
|
|
|
м-РНК |
|||||
|
|
|
|
|
|
|
|
|
|
|
м-РНК |
неактивный |
|
|
||
ИНДУКТОР |
репрессор |
|
трансляция |
||
|
лактаза |
|
пермеаза лактозы |
|
трансацетилаза |
|
|
|
|
|
Lac-оперон E. coli содержит 3 гена, отвечающие за образование белков, участвующих в переносе в клетку дисахарида лактозы и в ее расщеплении.
Z - - галактозидаза (лактаза) -расщепляет лактозу на глюкозу и галактозу. Y- - галактозидпермеаза - переносит лактозу через мембрану клетки.
А - тиогалактозидтрансацетилаза - ацетилирует галактозу.
144

Задания для самопроверки и самокоррекции
Задание 1 Для диагностики ВИЧ-инфекции используется метод полимеразной цепной реакции. Что лежит в его основе?
A.Амплификация генов
B.Рекомбинация генов
C.Транскрипция РНК
D.Сплайсинг РНК
E.Элонгация трансляции
Задание 2 При отравлении ядом бледной поганки блокируется работа РНК-полимеразы В ( ІІ). Какой процесс при этом нарушается?
A.Обратная транскрипция
B.Синтез праймеров
C.Синтез мРНК
D.Синтез тРНК
E.Созревание РНК
Задание 3 Геном человека содержит более 30 тысяч генов, а количество вариантов антител достигает миллионов. Благодаря какому механизму это возможно?
А. Образованию фрагментов Оказаки
B.Рекомбинации генов
C.Амплификации генов
D.Репарации ДНК
E.Репликации ДНК
Задание 4 Процесс биосинтеза белка является мишенью для многих антибиотиков. Какой процесс из
нижеперечисленных блокирует тетрациклин?
A.Элонгацию трансляции
B.Терминацию репликации
C.Терминацию трансляции
D.Элонгацию транскрипции
E.Инициацию репликации
Задание 5 При пигментной ксеродерме кожа очень чувствительна к солнечному свету. Дефект какого фермента
имеет место при данном заболевании?
A.ДНК-геликазы
B.Праймазы
C.УФ-эндонуклеазы
D.ДНК-гликозидазы
E.5’, 3’ - экзонуклеазы
Правильность решения проверьте, сопоставив их с эталонами ответов. |
|
1-A, 3-B, 5-C. |
146 |

РАЗДЕЛ 9. ВВЕДЕНИЕ В
ЭНДОКРИНОЛОГИЮ
Для координации многочисленных специфических функций в дифференцированных тканях многоклеточных организмов необходимы механизмы межклеточной регуляции.
В механизмах регуляции можно выделить три уровня:
Внутриклеточный уровень - осуществляется за счет ферментов (изменения их активности или количества)
Межорганную регуляцию выполняют две основные системы:
эндокринная система (второй уровень) - совокупность всех эндокринных желез, использующая в качестве мобильных посредников гормоны;
нервная система (третий уровень) |
- система проведения сигналов, обладающая |
|||||||
жесткой структурой. |
|
|
|
|
|
|
|
|
Все три уровня теснейшим образом связаны и функционируют как единая система, |
||||||||
накладываясь и перекрываясь. |
|
|
|
|
В иерархии регуляторных систем |
|||
|
|
|
|
|
|
|||
|
ЦНС |
|
|
|
|
главенствующая роль принадлежит |
||
|
|
|
|
|
нервной системе , которая получает |
|||
|
предшест- |
|
|
|
|
сигналы как внешней, так и внутренней |
||
|
|
|
|
|
среды, в результате чего возникают |
|||
|
венники |
|
|
|
|
|||
гипоталамус |
|
|
|
|
|
импульсы в специ-ализированных |
||
|
|
|
|
|
рецепторах. Импульсы поступают в |
|||
|
|
|
|
|
|
гипоталамус, где синтезируются |
||
либерины |
|
статины |
первые биологически активные |
|||||
|
гормональные вещества |
- |
рилизинг- |
|||||
|
|
|
|
|
|
факторы . Они не поступают в общий |
||
|
предшест- |
|
|
|
|
ток крови, а по системе портальных |
||
аденогипофиз |
венники |
|
|
|
|
сосудов достигают |
|
|
|
|
|
|
|
|
гипофиза |
||
|
|
|
|
|
|
специализированных клеток |
||
|
|
|
|
|
|
и оказывают стимулирующее |
|
|
|
|
|
|
|
отрицательная |
(либерины) или тормозящее |
|
|
тропные гормоны |
обратная |
(статины) действие на биосинтез и |
||||||
железы |
связь |
железу, вызывая секрецию |
|
|||||
|
|
|
|
|
|
выделение тропных гормонов. Эти |
||
|
предшест- |
|
|
|
|
гормоны с током крови поступают в |
||
периферические |
венники |
|
|
|
|
соответствующую |
эндокринную |
|
|
|
|
|
|
||||
|
|
|
|
|
|
определенного гормона. Гормоны |
||
|
|
|
|
|
|
действуют на органы или |
|
ткани- |
гормоны |
|
|
|
|
мишени, изменяя в них концентрацию |
|||
|
|
|
|
|
|
метаболитов, т.е. влияя на обмен |
||
|
|
|
|
|
|
веществ. Избыток гормона по |
|
|
клетки- |
|
|
|
|
|
принципу обратной отрицательной |
||
мишени |
ответ клетки |
|
|
|
|
связи тормозит выработку гормонов |
||
|
|
|
|
|
гипоталамуса или гипофиза. |
146 |
||
|
|
|
|
|
|
|||

Гормоны - (hormao -греч. - побуждать к действию) - вещества органической природы, вырабатываемые в специализированных клетках желез внутренней секреции, поступающие непосредственно в кровь и оказывающее регулирующее влияние на обмен веществ в тканях-мишенях и на функции всего организма.
Каждый гормон, где бы он ни был синтезирован, попадает в кровь и циркулирует вместе с ней, встречаясь на своем пути с клетками всевозможных типов. Но далеко не все клетки являются мишенями для любого гормона, они реагируют на присутствие в крови лишь одного или нескольких, но тоже строго определенных гормонов.
|
межклеточное |
гормон |
пространство |
|
|
кровь
рецептор
клетка- |
клетка - |
|
немишень |
||
мишень |
||
|
При этом все 75 триллионов клеток организма человека являются мишенями для одного или нескольких из 50 известных гормонов.
Реакция ткани-мишени на действие гормонов зависит от ряда факторов:
скорости продуцирования гормона в кровь и скорости деградации его в печени или в почках (от нескольких минут до нескольких суток);
анатомической близости ткани-мишени к источнику (эндокринной железе); скорости ассоциации и диссоциации гормонов со специальными белкамипереносчиками. Гормоны находятся в крови как в свободном, так и в связанном
состоянии - с протеинами плазмы, специфическими белками-переносчиками (резервуар гормонов, их неактивная форма). В тканях-мишенях гормон освобождаются от белка и начинают функционировать. В клинике определяют свободную и связанную формы
(например, повышение свободных женских половых гормонов |
- плохой |
диагностический признак). |
|
Все гормоны взаимодействуют с клетками-мишенями через |
рецепторы - особые |
молекулы узнавания, специфические белки сложной, уникальной структуры, обеспечивающие высочайшую избирательность. Один гормон может иметь несколько рецепторов. Нарушение функции рецепторов может лежать в основе многих эндокринных заболеваний. Нарушения могут быть следующие:
появление антител против рецепторов определенных гормонов; |
|
отсутствие связывания гормона с рецептором; |
|
нарушение регуляции рецепторов. |
148 |

Истинные гормоны обладают следующими отличительными свойствами: целенаправленность и дистантность действия (образуются в одних органах,
действуют в других); высокая биологическая активность (концентрация гормонов в крови ничтожно мала
(10-7 - 10-12 моль/л);
высокая специфичность (гормоны действуют только на свои ткани-мишени). Кроме истинных гормонов, секретирующихся непосредственно в кровь и обладающих дистантностью, существуют гуморальные регуляторные факторы
неэндокринного происхождения ( гормоны локального действия |
), регулирующие |
|
|||||||||
метаболизм там, где они вырабатываются ( |
изокринное действие ). Они образуются в |
|
|||||||||
разных органах и тканях: лимфоидной системе, лейкоцитах, соединительной ткани, |
|
|
|||||||||
ЖКТ, нервной системе, почках, миокарде, поэтому еще называются |
|
|
тканевыми |
||||||||
гормонами или гормоноидами. |
|
|
лютропин |
|
импульсы |
|
|||||
|
Концентрация гормонов подвержена |
|
30 |
|
|
||||||
|
|
|
|
|
|
|
|
|
|||
периодическим колебаниям, которые могут зависеть |
20 |
|
|
|
|
|
|
|
|
||
от времени года, времени дня или менструального |
10 |
|
|
|
|
|
|
|
|
||
цикла. Многие гормоны поступают в кровь |
|
|
|
|
|
|
|
|
|
||
|
|
12 |
15 |
18 |
21 |
24 |
3 |
6 |
9 |
||
импульсами и нерегулярно, их концентрация может |
|
||||||||||
|
|
|
время, сут |
|
|
|
|||||
пульсировать. Концентрация другой группы |
|
|
|
|
|
внешнее |
|
|
|||
гормонов меняется в зависимости от |
внешних |
30 |
|
|
|
|
|
|
|||
|
|
|
|
воздействие |
|||||||
факторов (стресс, голодание, прием пищи). Выброс |
20 |
инсулин |
|
|
|||||||
|
|
|
|
|
|
||||||
гормонов является ответом организма на внешнее |
|
|
|
|
|
|
|
||||
10 прием пищи |
|
|
|
|
|
||||||
воздействие или на изменение внутреннего |
|
|
12 |
15 |
18 |
21 |
24 |
3 |
6 |
9 |
|
состояния. |
|
|
|
|
время, сут |
|
|
|
|||
I. Классификация гормонов
Существует несколько подходов к классификации гормонов:
по растворимости в воде и липидах (гидрофильные или липофильные гормоны); по локализации рецепторов гормонов (мембранные или цитозольные рецепторы); по секретирующим эндокринным железам (гормоны гипофиза, поджелудочной,
щитовидной железы и т.д.); по метаболическому эффекту (гормоны гипергликемические, анаболические и т.д.)
по химической структуре:
гормоны белково-пептидной природы:
сложные белки (чаще гликопротеины): СГ, ТТГ;
простые белки: инсулин, СТГ;
пептиды: кальцитонин, глюкагон, нейропептиды, тканевые биорегуляторы; гормоны - производные аминокислот: тироксин, адреналин, норадреналин и др.; гормоны стероидной природы : кортикостероиды и половые гормоны,
производные витамина D;
эйкозаноиды – гормоноподобные вещества - производные арахидоновой кислоты: 149 простагландины, простациклины, тромбоксаны, лейкотриены.