

книги из ГПНТБ / Юдович, Ю. Б. Промысловая разведка рыбы учебник
.pdfир.макс = 25/р см/с (при длине рыбы / < 1,5 см),
з _
°р.макс = '25 у11 см/с (при длине рыбы I > 150 см).
Промежуточные значения могут быть получены по графику
(рис. 13).
Экспериментальные данные хорошо совпадают с теоретиче скими расчетами. При этом оказывается, что отношение макси мальной скорости к скорости миграции у рыб данной длины примерно постоянно.
Длина рыб, см |
12 |
20 |
50 |
100 |
200 |
Скорость максимальная, |
см/с 110 |
190 375 |
530 |
700 |
|
Скорость миграции, см/с |
45 |
53 |
75 |
117 |
140 |
Для промысловой разведки важно знать скорость переме щения косяков рыбы. Эти скорости определяют приближенно с помощью, например, судового лага, уравнивая скорость судна, идущего параллельно косяку, со скоростью косяка. Так была определена максимальная скорость косяков дальневосточной скумбрии во время их отхода из районов нагула на зимовку. Она составила 6 узлов. Скорость косяков сайры — 2—2,5 узла. Скорость косяков сахалинской сельди за 1—2 дня до нереста при испуге доходит до 4—5 узлов. В. Н. Гиренко приводит дан ные о еще больших скоростях нагульной западнокамчатской сельди.
По данным В. Н. Войниканиса-Мирского и Ю. Ю, Марти, ко сяки пеламиды двигаются со скоростью 3,2—5,0 узлов, сарди ны— до 2,2 узла и хамсы— 1,2—1,6 узла.
Рис. |
13. Кривые зависимости |
максимальной |
скорости |
(Л) |
и скорости 'миграций (В) |
рыб, дельфинов |
и китов |
|
от длины тела (по Шулейкину В. В.). |
||
43
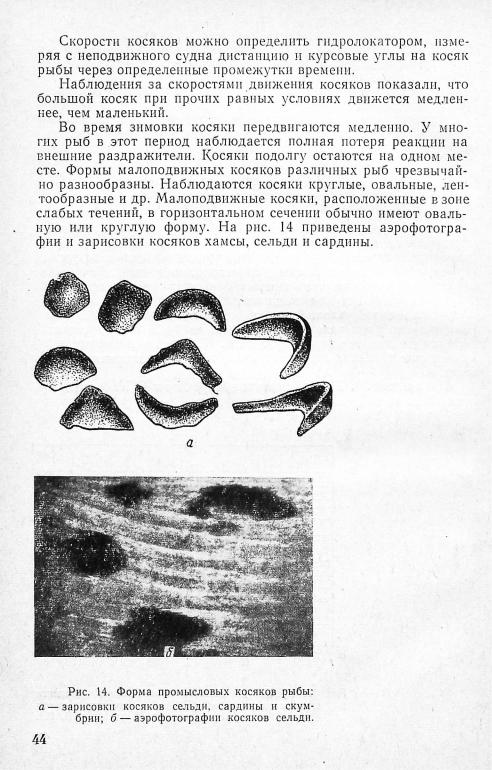
Наблюдения за формой движущихся косяков подтверждают е ы в о д ы академика В. В. Шулейкина о влиянии гидродинамиче ских законов на форму построения рыб при их движении: ми нимальное сопротивление воды стая крупных рыб будет испы тывать при движении «журавлиным» строем или строем «усту па». Такой строй наблюдается у дельфинов, косаток, тунцов. Движущиеся косяки сельди, сардины имеют серпообразную форму, причем в вершине «серпа» рыба держится более плотно. Мелкие рыбы при движении образуют косяки каплевидной фор мы. Это, по мнению В. В. Шулейкина, объясняется тем, что плотная стая рыб обтекается примерно так же, как сплошное тело, и, следовательно, при каплевидной форме сопротивление
минимально.
По характеру эхограммы движущийся косяк можно легко отличить от малоподвижного. В первом случае он имеет чет кий контур и одну вершину. Малоподвижные косяки имеют «размытые» контуры и несколько вершин (см. рис. 8).
МИГРАЦИИ
Большинство рыб, китов, крабов и других промысловых объектов совершают миграции — регулярные перемещения из одного района в другой. Различают горизонтальные и верти кальные миграции. Сезонные горизонтальные миграции многих рыб и других водных животных — явление столь же постоянное, как перелеты птиц. К сезонным миграциям относятся нагульные (перемещение к местам нагула), нерестовые (перемещение к местам нереста) и зимовальные (движение к местам зимовки). Сезонные миграции связаны с тем, что в разные периоды жизни рыбы нуждаются в различных условиях среды, например осе тровые нагуливаются на морских мелководьях, богатых бенто сом, а на нерест входят в реки. Океанические сельди нагули ваются вдали от берегов, над большими.глубинами, а на нерест подходят к берегам или на банки.
Кормовые миграции связаны с распределением и количест вом пищи, температурными условиями. По мере выедания корма скопления перемещаются к новым кормовым полям-.
Нерест рыб происходит в определенных местах, где обеспе чены хорошие условия для развития икры и выживания маль ков. Одни рыбы находят такие условия вблизи берегов, дру гие— в открытых водах. Сроки начала и конца миграций могут отклоняться от средних многолетних значений на 15—20 суток, а иногда и более. Они зависят от температуры, течений, режима ветра, распределения пищи. Так, неблагоприятные условия среды на нерестилищах (подход льда, сильное охлаждение воды) могут задержать сроки нереста. Средняя скорость мигра ции зависит от характера миграции, а также от условий среды. Она колеблется от 1—2 до 20—25 миль в сутки. Результаты
меченая показали, что тунцы пересекают Тихни и Атлантиче ский океаны со средней скоростью 8—9 миль в сутки. Скорость нерестовой миграции в основном определяется степенью зрело сти половых продуктов, расстоянием до нерестилищ и гидроло гическим режимом. Скорость кормовой миграции -зависит от ко личества пищи и темпов изменения температуры, а зимоваль ной— от упитанности рыбы и темпов понижения температуры воды. Протяженность миграций у разных объектов очень раз нообразна. Тунцовые и мечеобразные мигрируют на расстояния 6—7 тыс. миль, осетровые — на 1—1,5 тыс. миль, тресковые и некоторые океанические сельди — на 500—800 миль, камбала и хамса — на 200—300 миль, тюлька и бычки — на 100—150 миль.
Направления, пути и сроки миграций многих промысловых объектов хорошо изучены. Часто промысел рыбы существует в течение всего года, когда рыбу ловят не только на нерестили щах и зимовках, но и на миграционных путях.
Важно изучать вертикальные миграции, которые совершают почти все пелагические и донные рыбы. От особенностей верти кального распределения рыбы зависит выбор орудий лова, тех ники и организации промысла. Эти миграции бывают сезонные, возрастные и суточные. Сезонные вертикальные миграции — это переход с глубоких участков водоема на более мелкие или об ратно. Такие миграции характерны для донных объектов — кам бал, палтусов, окуней, макрурусов, крабов. Эти миграции харак терны медленной сменой глубин. Для промысловиков особенно важно знать суточные вертикальные миграции объекта. Почти все рыбы-планктонофаги (сельдь, скумбрия, сардина, хамса, килька и др.) регулярно совершают суточные вертикальные ми грации. Амплитуда этих миграций бывает весьма значительной. Б. Г1. Мантейфель, С. Г. Зуссер и другие ученые, исследовавшие вертикальные миграции рыб, считают, что они имеют приспосо бительное значение. Так, у рыб-планктонофагов северных морей отношение к факторам среды меняется по сезонам. Оно связано с особенностями изменения кормовой базы. Обилие пищи наб людается 2—3 летних месяца. В это время планктонофаги от кармливаются, а остальную часть года питаются слабо или вообще не питаются, расходуя накопленный жир. Следователь но, в период откорма их вертикальные миграции подчинены задаче наиболее интенсивного питания.
Вертикальные миграции большую часть года прямо или кос венно связаны с изменением освещенности толщи воды, поэтому на судах промысловой разведки организованы регулярные на блюдения за вертикальным перемещением рыб в связи с осве щенностью. Обычно в светлое время суток рыба держится глуб же, чем ночыо. В сумерки начинается миграция к поверхности.
Ритм изменения освещенности настолько точно совпадает с ритмом суточных вертикальных миграций, что может служить показателем глубины нахождения косяков рыбы.
46
Анализ большого фактического материала, проведенный С. Г. Зуссер (ВНИРО), показывает, что вертикальная миграция рыб, как правило, зависит от сроков смены освещенности и еже суточный подъем рыбы к поверхности совпадает с изменением освещенности воды. Распределение температуры, солености, кислорода и других факторов не оказывают существенного вли яния на характер суточных вертикальных миграций, если пара метры этих факторов ие достигают величин, при которых оби тание рыбы становится невозможным. Например, норвежская весенненерестующая сельдь в декабре ежесуточно опускается ут ром на глубину, где температура близка к нулю, а вечером под нимается к поверхности и держится при температуре 4,5° С. Суточные вертикальные миграции этой сельди не прекращаются и при уменьшении кислорода на глубине, где сельдь держится днем. Вечерний подъем планктоноядных рыб происходит в ве черние сумерки по мере уменьшения освещенности. Примерная скорость подъема составляет 0,5—2,5 см/с. В бореальной обла сти большинство планктонофагов мигрируют днем на глубину 300—600 м, а ночью поднимаются и держатся в 10—100 м от поверхности. В тропической области амплитуда миграций боль шинства рыб шельфовой зоны не превышает 150—200 м, а в тро пиках южного полушария, где преобладают рыбы, питающиеся фитопланктоном, амплитуда их вертикальных миграций состав ляет всего 10—15 м.
Скорость вертикальных миграций планктоноядных рыб не за висит от длины особей и видовых особенностей рыб. Наиболее выражены суточные вертикальные миграции у планктоноядных рыб. С. Г. Зуссер подсчитывала число косяков планктоноядных рыб на различной глубине; оказалось, что наибольшее число ко сяков встречается на глубине, где освещенность составляет 0,1— 0,01 лк. Это подтверждается и гистологическими исследования ми— процесс темповой адаптации глаза, переход к более чувст вительному палочковому зрению завершается при освещенности около 0,1 лк.
Изучение особенностей вертикальных миграций в связи с факторами внешней среды способствует установлению новых закономерностей п совершенствованию поисковой работы.
ПРОМЫСЛОВЫЕ И НЕПРОМЫСЛОВЫЕ ОБЪЕКТЫ
Промысловыми объектами принято называть рыб или дру гих животных, для лова которых существует специальный про мысел. К промысловым объектам относятся сельдь, треска, оку ни, палтусы, скумбрия, тунцы и др. К непромысловым объектам относятся луна-рыба, некоторые виды окуней, бычков и др. Важная особенность всех промысловых объектов — их высокая численность, обеспечивающая организацию специализированно
47
го промысла. Поэтому в разных районах рыболовства одни н те же виды рыб могут относиться как к промысловым, так и непромысловым объектам. Например, в Баренцевом море тре ска является одним из основных промысловых объектов, а в юго-восточной части Берингова моря она ловится единично и является так называемым «приловом» — непромысловым объек том. Бычки в Азовском море — важный промысловый объект, а у берегов Сахалина их не ловят. Некоторые рыбы, даже обла дающие высокой численностью, не считаются промысловыми. Чем выше потребность в рыбных продуктах, тем больше объек тов становится промысловыми. Понятия «промысловый» и «пепромысловый» объект тесно связаны с техническим прогрессом в области промышленного рыболовства, судостроения, рыбооб рабатывающего машиностроения, технологии переработки сырца.
Таким образом, понятие промысловый объект меняется с течением времени. Объекты, лов которых не был организован, начинают облавливаться промысловым флотом и становятся промысловыми. Те объекты, которых начали облавливать, ред ко переходят в разряд непромысловых, однако промысел может уменьшаться пли прекращаться вообще с падением их числен
ности. В таких случаях объект теряет |
промысловое значение |
и становится непромысловым (крабы у |
восточного побережья |
Камчатки, пеламида в Черном море). Многие объекты, являю щиеся непромысловыми для одного государства, представляют большой промысловый интерес для другого (иглокожие, неко торые виды рыб, земноводные).
Каспийские кильки долгие годы считались непромысловым объектом и лишь с разработкой техники их лова с помощью электросвета эта рыба стала промысловой, дающей свыше 70% вылова в Каспийском море. Добыча морских окуней стала воз можной с постройкой траулеров. Крабы стали промысловым объектом после разработки технологии их консервирования
ит. д.
Сразвитием техники рыбообработки появляется возмож
ность использования рыб, ранее считавшихся непромысловыми, и других непромысловых в настоящее время животных (кра- бов-стригунов и др.).
Промысловые объекты делятся на пищевые и непищевые. Первых используют для выработки продуктов питания, вторых для технических нужд — производства рыбной муки, кормов для скота, выработки технического жира. Некоторые объекты используют для выработки и пищевой и технической продукции (анчоус, сельдь, килька).
Так как непромысловые рыбы могут переходить в разряд промысловых, во время поисков и разведки необходимо учиты вать и изучать все объекты, встречающиеся в районе-работы эк спедиций.
48
КОЛИЧЕСТВЕННАЯ ОЦЕНКА ПРОМЫСЛОВЫХ КОСЯКОВ И СКОПЛЕНИЙ
Количественная оценка промысловых скоплений необходима для того, чтобы определить их размеры и подсчитать величину возможного вылова, характеризующую степень возможного ис пользования скопления в данный период. Способы оценки дон ных, придонных и пелагических скоплений различны, но во всех случаях для получения количественной оценки скопление оконтуривают. Для этого в зависимости от типа скопления его пере секают либо двумя поисковыми галсами («крестом»), либо сетью поисковых галсов (рис. 15). При оконтуриваиии судно движется по разрезу до тех пор, пока гидроакустические прибо ры фиксируют рыбу пли пока орудия лова улавливают ее. Кон тур скопления наносят на планшет по данным крайних наблю дений. Когда его наносят по данным об уловах, предполагают, что улов плавно изменяется от одного пункта к другому. Про должительность контрольных ловов, орудие лова и режим рабо ты при оконтуриваиии скопления должны быть неизменными. Если продолжительность лова не равна принятому эталону, то величину улова приводят в соответствии с принятой продолжи тельностью облова.
|
скоплений: |
а — «крестом»;- |
б — серией поисковых галсов; 1 — про |
мысловый улов, |
2 — непромысловый улов, 3 — улов от |
|
сутствует. |
|
ДОННЫЕ СКОПЛЕНИЯ |
Количественную оценку донных скоплений получают по конт рольным обловам тралами, ставными сетями или донными яру сами. Наиболее распространенный способ контрольного лова донных скоплений — траловая съемка. Траления считаются со поставимыми, если они получены в периоды с однообразным
49
поведением объекта поисков. Если в течение суток улов изме няется закономерно, то время траления выбирается с учетом этих изменений. Для оценки возможного улова подсчитывают его величину в пределах контура скопления.
Предполагая, что рыба распределена по площади траления равномерно, величину возможного вылоеэ на скоплении Q можно определять по формуле
Q = - у (ч ± е),
где F — оконтуренная площадь скопления;
/ — площадь одного контрольного траления;
q — средний улов за траление;
е — погрешность в определении.
Если в оконтуренном скоплении уловы отличаются значи тельно, следует проводить вспомогательные контуры, исполь зуя точки равных уловов и выбирая их так, чтобы между этими контурами улов изменялся не более чем в 2—3 раза. Величину возможного вылова следует определять суммированием величин возможных выловов на площадях F\, F2, ..., Fn, заключенных между промежуточными контурами.
Q |
= |
~ |
(<7l ± ei ) + |
- у ~ (<?2 ± |
ез ) т |
• • |
• Н— j ~ |
( Я п |
± ел ) |
= |
|
|
|
|
= | ] Д |
й |
± е,). |
|
|
|
|
|
|
|
|
|
|
|
|
|
Т А Б Л И Ц А 4 |
|
Номер |
|
|
q — q (от- |
|
Номер |
|
q — q |
(от- |
|
|
контроль |
|
<7, ц |
клоненне |
|
контроль |
|
клоление |
|
||
ного тра |
|
от сред |
( q — q ) 2 |
ного тра |
|
от сред |
( q — q ) 2 |
|||
ления |
|
|
него) |
|
ления |
|
него) |
|
||
1 |
• |
13 |
+ 6 |
36 |
|
8 |
12 |
+ 5 |
25 |
|
2 |
|
7 |
0 |
0 |
|
9 |
8 |
+ 1 |
1 |
|
3 |
|
5 |
_2 |
4 |
|
10 |
7 |
|
0 |
0 |
4 |
|
7 |
0 |
0 |
|
11 |
3 |
— 4 |
16 |
|
5 |
|
7 |
0 |
0 |
|
12 |
■ 7 |
|
0 |
0 |
6 |
|
4 |
—3 |
9 |
|
13 |
5 |
—2 |
4 |
|
7 |
|
6 |
—1 |
1 |
|
|
|
|
|
|
|
|
|
|
|
|
|
Б ? = |
|
|
2 ( q - qf = |
|
|
|
|
|
|
|
=91 |
|
|
=96 |
50
Среднее квадратическое отклонение а величины улова за траление qi от среднего значения q определяется по формуле
а =
2 (дс — д)-
п — 1
где п — число тралений.
Для определения величины возможного улова данные о контрольных уловах заносят в таблицу (табл. 4) и по ней на ходят средний улов д, среднее квадратическое отклонение о и коэффициент вариации уловов.
9 = |
|
91_ |
п |
= 7 ц, |
|
|
13 |
|
а - - |
|
96 |
|
2,8 ц, |
|
|
|
12 |
С |
|
4096. |
Величину ошибки е определяют по формуле
s = /п |
0 |
/— , |
|
1 у |
п |
где ip— коэффициент, зависящий от доверительной вероятности и числа кон
|
трольных тралений без единицы (табл. 5). |
|
|
|
|
|||||
|
|
|
|
► |
|
|
ТАБЛИЦА s |
|||
|
|
Значения t 0 |
при р |
|
|
|
Значения |
iQпри (5 |
||
/1—1 |
|
Р |
|
|
п-1 |
|
|
Р |
|
|
0,7 |
0,8 |
0,9 |
0,95 |
0.7 |
0,8 |
0,9 |
0.95 |
|||
|
|
|||||||||
8 |
1,108 |
1,397 |
1,860 |
2,31 |
12 |
1,079 |
1,356 |
1,772 |
2,18 |
|
9 |
1,100 |
1,383 |
1,833 |
2,26 |
15 |
1,074 |
1,341 |
1,753 |
2,13 |
|
10 |
1,093 |
1,372 |
1,812 |
2,23 |
20 |
1,064 |
1,325 |
1,725 |
2,09 |
|
11 |
1,088 |
1,363 |
1,796 |
2,20 |
|
|
|
|
|
|
В практике промразведки |
|3 |
следует принимать равной |
|
0,8—0,9. |
|
|
(при р= 0,9, п—1 = 12) |
Для приведенного выше примера |
|||
tp = 1,77, |
|
|
|
s = 1,77 |
= |
± |
1,4 ц. |
Величина улова за траление |
с вероятностью 0,9 составляет |
||
9 = 7 + |
1,4 ц. |
|
|
Предположив, что — =0,0001, находим величину возмож-
F
иого вылова на скоплении
Q = 70 000 ± 14 000 ц.
51
В практике промысловой разведки для предварительной оценки района погрешность следует принимать со знаком ми нус, с тем чтобы не завысить величину скопления. Указанное обстоятельство чрезвычайно важно, так как погрешность может достигать большой величины. Поэтому величина возможного улова на скоплении должна быть определена 56 000 ц.
После оконтуривания скопления на планшете его площадь можно определить планиметром, палеткой или аналитическим вычислением. В последнем случае оконтуренное скопление раз бивают на геометрические фигуры, площадь которых можно вычислить по элементарным формулам. Вычисление планимет ром следует выполнять дважды. Разница между измерениями не должна превышать 5%. Для расчетов принимают среднюю величину. Измерительная палетка — лист кальки, разделенный на клетки, в центре которых проставлены точки. Палетку накла дывают в нескольких произвольных положениях на оконтурен ную площадь, подсчитывают число точек, попавших на эту пло щадь, и умножают это число на величину площади клетки па летки, деленную на масштаб планшета. Среднюю величину из 3—5 подсчетов принимают за искомую. Применение палетки дает хорошие результаты при определении больших площадей.
После определения величины возможного улова в ряде слу чаев приходится учитывать снижение уловов при облове скоп ления промысловым флотом. Инженер Запрыбпромразведкп К- Г. Кухоренко предложил метод определения оптимального количества траулеров в районе промысла, при котором величи на улова на траление не снижается. В качестве критерия насы щенности флотом К. Г. Кухоренко принял отношение площади, протраленной за сутки всеми судами, к площади, занимаемой скоплением. Им было установлено, что па промысле скумбрии донными тралами предельная величина этого отношения состав ляет 0,5—0,8. Увеличение отношения приводило к снижению улова. Очевидно, что величина коэффициента вылова в разных районах может варьировать. Она зависит от интенсивности под хода новых скоплений, уловистости тралов и других факторов.
Часто не вся площадь, занятая скоплением, может представ лять промысловый интерес, так как зоны высоких уловов чере дуются с зонами низких непромысловых уловов. В подобных случаях величину возможного вылова следует определять для зон с промысловой плотностью рыбы.
Размеры скоплений не остаются постоянными во времени, поэтому подобные вычисления делают несколько раз (й разные периоды) и сравнивают результаты. Часто эти результаты силь но отличаются один от другого. Эти различия могут произойти за счет изменения уловистости тралов в течение года. Чем ме нее подвижны рыбы, чем меньше они реагируют на приближе ние трала, тем больше уловистость. Учитывая это, наиболее до стоверными результатами следует считать те, которые получены
.52