

3.3. Самоідентифікація та взаємне упізнавання бактерій.
У ході еволюції в різних організмів розвилися різноманітні, у тому числі досить складні й ефективні, механізми розрізнення «свого-чужого». Здатність відрізняти своїх від чужих – одна з фундаментальних властивостей живого. На цій здатності засновані найважливіші біологічні процеси і явища, у тому числі статеве розмноження (вибір партнера), видоутворення (формування репродуктивної ізоляції), захист від патогенних мікробів і вірусів, відторгнення чужорідних тканин, колоніальність, багатоклітинність, соціальність, включаючи різні форми кооперації й навіть альтруїзму (який, як правило, спрямований на «своїх», насамперед на родичів).
У вищих організмів (еукаріот) ці механізми вивчені значно краще, ніж у прокаріот (бактерій і архей).
Оскільки генетичні й біохімічні основи розрізнення «своїх» і «чужих» у бактерій вивчені поки недостатньо, кожне нове відкриття в цій області притягає до себе пильну увага.
Американськими мікробіологами був розкритий механізм взаємної ідентифікації бактерії протея (Proteus mirabilis) (Рис.3.6). Цей мікроб мешкає в травному тракті людини й зазвичай не є шкідливим, але при несприятливому збігу обставин може викликати інфекційні захворювання.
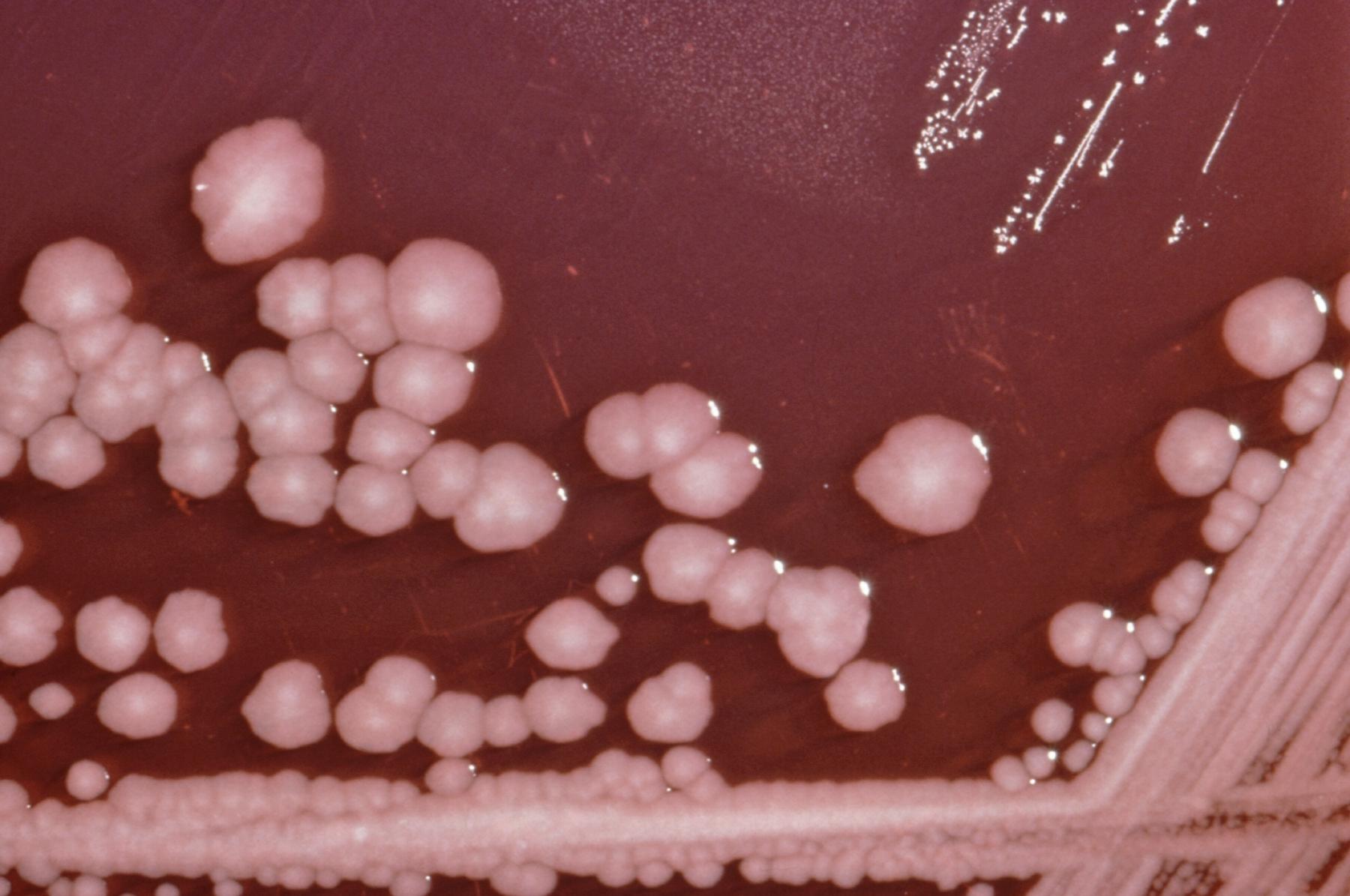
Рис.3.6. Бактерії Proteus mirabilis
Для протея характерний досить складний аналог соціальної поведінки, що виражається, зокрема, у погоджених пересуваннях великих груп бактеріальних клітин.
У геномі цієї бактерії виявлений ділянка, відповідальна за самоідентифікацію й взаємне упізнавання мікробів. Мутації в цій ділянці можуть приводити до миттєвого поділу популяції мікробів на групи, що ставляться одна до одної як до «чужинців». Представники таких груп «відмовляються» утворювати змішані скупчення.
Коли дві колонії або «зграї» (swarms) Proteus mirabilis зустрічаються на поверхні живильного, середовища відбувається одне з двох: або колонії зливаються, або вони зберігають самостійність. В останньому випадку між ними утвориться добре видима межа. Вибір одного із двох варіантів залежить від ступеня спорідненості. Якщо обидві зграї належать до того самого штаму (клону, генетичної лінії), вони, швидше за все, зіллються; якщо до різних –між ними, швидше за все, виникне границя. Дана особливість використовується в медичній практиці для ідентифікації штамів мікроба.
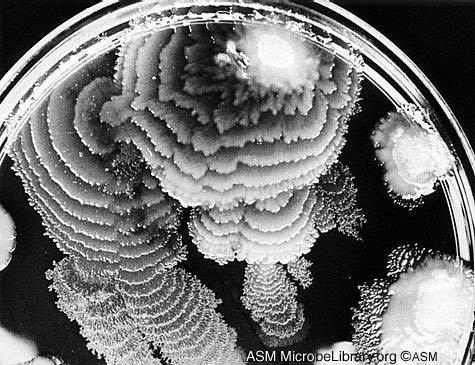
Рис.3.7. Колонії Proteus mirabilis
Було встановлено, що багато штамів Proteus mirabilis виділяють токсичні білки – протіцини, призначені для боротьби з конкуруючими штамами. Кожний штам виділяє власний протіцин, до якого він стійкий, і до того ж кожен штам має стійкість до деяких (але не до всіх) чужих протіцинів. Однак утворення меж між неспорідненими «зграями» обумовлюється не протіцинами, а чимось іншим. Справа в тому, що деякі штами не виділяють протіцинів, але при цьому однаково утворять межі, у тому числі й з іншими штамами, позбавленими протіцинів.
Щоб з'ясувати причини цього явища, дослідники «змусили» бактерій швидко мутувати, активізувавши переміщення мобільних генетичних елементів (транспозонів). З одного «предкового» штаму (BB2000) було отримано 3600 мутантних штамів-нащадків. Серед них виявився один, котрий «відмовився» змішуватися з іншими мутантами та із предковим штамом BB2000. Його назвали Ids (від слів «identification of self» – «самоідентифікація»). Очевидно, у штамі Ids відбулася мутація, внаслідок якої цей штам змінив свою самоідентифікацію й почав сприймати батьківський штам BB2000 як «чужий».
Виявилось, що в штамі Ids транспозон вмонтувався в ділянку генома, що містить шість генів, функції яких дотепер не були відомі. Учені позначили цю ділянку idsABCDEF. Очевидно, транспозон порушив нормальну роботу цих генів, що й призвело до «відчуження» бактерій-мутантів від предків і родичів.
Щоб перевірити це припущення, у бактерій із предкового штаму BB2000 видалили всі шість генів. Вийшли мутанти, які «вважали» штам Ids «своїм», а предків - «чужими». Стало ясно, що дані шість генів дійсно відіграють ключову роль у самоідентифікації цих бактерій і в розрізненні «своїх» і «чужих». Після цього почалася кропітка генно-інженерна робота з метою з'ясування функцій кожного із шести генів окремо.
З'ясувалося, що видалення гена idsA саме по собі не веде до зміни самоідентифікації бактерій. Цей ген, очевидно, грає якусь допоміжну роль. Однак видалення або поломка хоча б одного з генів idsB, idsC, idsD, idsE, idsF приводить до того, що бактерії починають вважати предковий штам «чужим». При цьому бактерії, у яких вилучений ген B, C, D або E, вважають «своїми» бактерій, у яких вилучені всі шість генів. Однак бактерії з вилученим геном F вважають «чужими» всіх, крім самих себе.
Дослідники виявили, що схожі генетичні системи ідентифікації є й в інших штамів Proteus mirabilis, а також в інших видів бактерій. Була проведена додаткова серія експериментів, у якій ділянку idsABCDEF пересаджували (цілком або вроздріб) іншому, неспорідненому штаму Proteus mirabilis (штам HI4320) на додаток до його власної генетичної ідентифікаційної системи. Бактерії зі штаму HI4320, яким ділянку idsABCDEF від штаму BB2000 пересадили цілком, починають вважати свій рідний штам HI4320 «чужим». Однак якщо в пересадженій ділянці ушкоджений ген D або E, відчуження не відбувається: бактерії продовжують вважати штам HI4320 «своїм».
Ці й інші, більш складні, експерименти дозволили вченим установити, що найважливішу роль у розрізненні «своїх» і «чужих» грають гени D і E. Саме вони визначають «індивідуальність» бактерії. У цих генах виявилися варіабельні області, тобто ділянки, що сильно різняться в різних штамів. Гени D і E утворюють унікальну генетичну «ідентифікаційну картку», відповідно до якої штами бактерій розрізняють один одного.
Гени B, C і F, очевидно, потрібні для того, щоб ця «ідентифікаційна картка» могла бути належним чином представлена й упізнана. У різних штамів ці гени різняться слабко, і якщо взяти кожен із цих генів в одного штаму й пересадити іншому, самоідентифікація бактерій не зміниться.
Судячи з ряду непрямих ознак, система ідентифікації в Proteus mirabilis не обмежена ділянкою idsABCDEF. Швидше за все, існують і інші, поки ще не виявлені, гени, що беруть участь у розрізненні «своїх» і «чужих». Вся система в цілому може виявитися досить складною.
Існують і інші ситуації, у яких бактерії можуть вирішувати дилему «свій»-«чужий». Наприклад, ситуація симбіозу.
Симбіоз із різними бактеріями надзвичайно широко розповсюджений як у тварин, так і в рослин. Дуже часто багатоклітинний організм не одержує симбіонтів автоматично від своїх батьків, а повинен сам якимсь чином знайти їх у навколишнім середовищі й «одержати» їх на початку свого життя. З огляду на величезну розмаїтість мікроорганізмів у всіх природних місцеперебуваннях, зрозуміло, що в такій ситуації першорядного значення набуває точність «взаємного упізнавання» симбіонта й хазяїна.
Але механізми цього дізнавання, безумовно, існують.
Є паразитичні бактерії, у яких дуже близькі форми паразитують на зовсім різних хазяїнах. Наприклад, один з різновидів бактерії Salmonella enterica заражає тільки людей, а інший має широке коло хазяїв – від приматів до гризунів. При цьому геноми обох різновидів надзвичайно схожі один на одного. Поки не вдалося виявити в їх геномах те ключове розходження, від якого залежить вибір хазяїна.
У випадку симбіозу обидва учасники зацікавлені в точнім дізнаванні партнера. Ученим вдалось розшифрувати механізм взаємного упізнавання в симбіотичній асоціації рослин з азотфіксуючими бактеріями. У цьому випадку ключову роль грають виділювані бактеріями сигнальні речовини, які певним чином впливають на роботу генів у клітинах кореня рослини-хазяїна.
Частково розшифрований і механізм вибору партнера в іншої пари симбіонтів – тихоокеанського кальмара Euprymna scolopes і бактерії, що світиться, Vibrio fischeri.

Рис.3.9. Кальмар Euprymna scolopes світиться завдяки симбіотичним бактеріям Vibrio fischeri, що живуть в особливому «світному органі».

Рис.3.10. Бактерії, що світяться, Vibrio fischeri
Відомо, що одні різновиди бактерії Vibrio fischeri мешкають у симбіозі з рибами, інші – з кальмарами. Учені з'ясували, що здатність заселяти світні органи кальмарів визначається одним-єдиним бактеріальним геном. Пересадження цього гена від «кальмарового» різновиду мікроба його «рибному» різновиду призводить до того, що останній набуває здатність жити в симбіозі з кальмарами.
Геноми «кальмарового» і «рибного» різновиду Vibrio fischeri були цілком вивчені й виявилися досить подібними (90% подібності за набором генів і 98,8% – за амінокислотними послідовностями білків).
Дослідники виявили, що один з тих генів, які активно працюють у Vibrio fischeri у період заселення світних органів молодих кальмарів, у геномі «рибного» штаму відсутній. Подальші дослідження показали, що цей ген (його символ – rscS) дійсно відіграє ключову роль у «співдружності» бактерій з кальмарами.
Ген rscS кодує білок-рецептор, що розташований на клітинній мембрані, він реагує на певні зовнішні сигнали й передає їх усередину клітини, активізуючи інший регуляторний білок – транскрипційний фактор SypG. У свою чергу, SypG стимулює активність групи генів, які кодують білки, необхідні для синтезу особливих полісахаридів, які виводяться з бактеріальної клітини й розташовуються на її зовнішній поверхні. Від цих полісахаридів залежить здатність бактерій розмножуватися в слизу, виділюваної світним органом кальмара.
Ген rscS зустрічається у двох алельних варіантах (rscSA і rscSB), причому тільки алель rscSA дозволяє бактерії вступити в симбіоз із кальмаром. Саме цей варіант гена присутній у всіх «кальмарових» штамів і в того «рибного» штаму, що зумів «налагодити стосунки» з кальмаром. У всіх інших «рибних» штамів або присутній інший варіант гена (rscSB), або ген зовсім відсутній (як у того «рибного» штаму, чий геном був повністю прочитаний ученими).
Аби переконатися в тому, що наявність алелю rscSA у V. fischeri є необхідною й достатньою умовою симбіозу з кальмарами, учені провели ряд генно-інженерних експериментів. Вони виводили з ладу цей ген в «кальмарових» штамів – і бактерії втрачали здатність жити в співдружності з кальмарами. Вони вставляли ген в «рибні» штами – і вони набували цю здатність. Таким чином, припущення цілком підтвердилося.
На основі порівняння нуклеотидних послідовностей трьох інших генів дослідники побудували еволюційне древо вивчених штамів мікроба. Судячи із цього древа, споконвічно V. fischeri не мав гена rscS і жив у симбіозі тільки з рибами. Потім один зі штамів мікроба набув цього гена. Всі штами, що мають ген rscS, утворюють на древі монофілетичну (що походить від одного предка) групу. Це означає, що ген був надбаний тільки один раз. Спочатку він існував тільки в одному варіанті – rscSA. Деякі з нащадків того мікроба, що вперше обзавівся новим геном, залишили своїх колишніх хазяїв – риб – і перейшли до симбіозу з кальмарами. Згодом дехто з них повернувся до симбіозу з рибами, причому це повернення супроводжувалося змінами в гені rscS. Так з'явився другий варіант цього гена – rscSB, що, можливо, якимсь чином поліпшує «взаєморозуміння» між мікробами й рибами.
Це перший відомий випадок, коли головною «діючою особою» у взаємнім упізнаванні симбіонтів виявився регуляторний ген.
Учені вважають, що необхідно подальше вивчення генетики процесів самоідентифікації й взаємного упізнавання бактерій для їх подальшого практичного використання.
Питання для повторення й обговорення.
1. Які елементи поведінкових реакцій можна вивчати у бактерій?
2. У чому виражається «колективне поведінку» у Myxococcus - типового представника міксобактерій?
3. Серед міксококків є генетичні лінії (або штами), не здатні до утворення власних плодових тіл, але вміють прилаштовуватися до «чужих» плодовим тілам і утворювати там свої суперечки. Як їм це вдається?
4. У результаті якої мутації бактерії Myxococcus xanthus придбали відразу два корисних властивості: здатність до складного колективного поведінки (утворенню плодових тіл) і захищеність від паразитів?
5. Що ви знаєте про генетику хемотаксису у бактерій?
6. Які гени рухливості і хемотаксису були встановлені на генетичної карті Е. coli?
7. За допомогою яких експериментів можна спостерігати хемотаксис у E. сoli?
8. Які виділяють категорії фенотипів мутацій хемотаксису у бактерій?
9. Які поведінкові реакції можуть проявляти бактерії?
10. Як здійснюється самоідентифікація та взаємопізнавання бактерій?