

6.7. Використання мозаїків для виявлення структур, що порушені при поведінкових мутаціях.
Відомо, що мозаїчні особини, які мають генотипічно різні частини тіла, являють величезну цінність для дослідження клітинної автономії, специфічності вираження генів у різних тканинах, взаємодії тканин різного генотипу й ін.
Характеристики мозаїків. Мозаїки — це складні особини, у яких деякі ділянки різних тканин можуть бути мутантними, у той час як інші мають нормальний генотип. Це дає можливість ідентифікувати ту частину, яка зачіпається дією мутантного гена при формуванні мутантного поведінкового фенотипу особини.
Статеві мозаїки (або гінандроморфи), в яких одні частини тіла мають генотип самця, а інші — самиці, можуть дати цікаву інформацію щодо організації статевої поведінки.
Перші роботи в цьому напрямку були проведені ще до 1932 року, коли Уайтинг вивчав статеву поведінку паразитичних ос (Habrobracon). У них відмінність у статевій поведінці не зводиться, як у дрозофіли, до відсутності в самиць реакцій, характерних для самців. Спостерігаючи гінандроморфів, які мають голову однієї статі, а іншу частину тіла — іншого, Уайтинг виявив, що поведінка визначається генотипом клітин головного ганглія. Білатеральні (ліво-праві) мозаїки виявляли тенденцію як до поведінки, характерної для самців, так і до поведінки, характерної для самиць.
Є кілька генетичних ситуацій, що ведуть до утворення гінандроморфів, а, отже, і кілька методів одержання мозаїків. Один з них заснований на використанні лінії дрозофіли з кільцевою X-хромосомою й залежить від втрати нестійкої кільцевої X-хромосоми під час першого поділу ядра зиготи ембріона самиці. У цьому випадку виникає два клони клітин: ХО й XX (Рис. 6.3.)
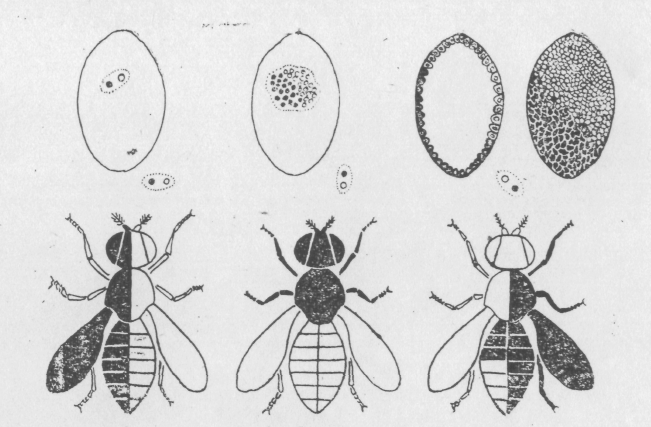
Рис.6.3. Соматична діаграма утворення генетичних мозаїків.
Співвідношення частин організму, що мають генотипи XX і ХО, залежить від того, коли втрачається кільцева X хромосома. Якщо, наприклад, вона втрачається при першому поділі ядра зиготи, то клітинні клони займають по половині бластодерми, якщо пізніше, то клон ХО займає меншу частину бластодерми. У рідких випадках кільцева X-хромосома втрачається при першому поділі й ще раз при більш пізніх поділах, у цих випадках в ембріонів клон ХО займає більшу частину бластодерми.
Характер же мозаїцизму буде визначатися не тільки віссю першого поділу, але й характером міграції ядер до поверхні при утворенні бластули. Відносне положення ядер має тенденцію зберігатися, але все-таки при міграції ядра з генотипом ХО можуть мігрувати разом з ядрами з генотипом XX.
Мігруючі ядра рівноспадкові, і при утворенні бластодерми місце, займане клітиною, в основному визначає її долю, тобто визначає ті структури, походження яким вона дасть. Цей механізм забезпечує появу різноманітних мозаїків.
Нервова система й мезодерма, що дає початок м'язовій тканині, також утворюються зі специфічних районів бластодерми. Границі в мозаїків між «чоловічими» і «жіночими» частинами в основному збігаються з міжсегментальними границями й поздовжньою середньою лінією екзоскелету (Рис. 6.4.), тому що екзоскелет представляє собою сукупність багатьох частин, що утворюються незалежно в ході метаморфоза з імагінальних дисків личинки. Імагінальні диски, у свою чергу, утворюються зі специфічних районів бластодерми. Однак окремі клітинні групи усередині одного диска можуть бути також різного генотипу.
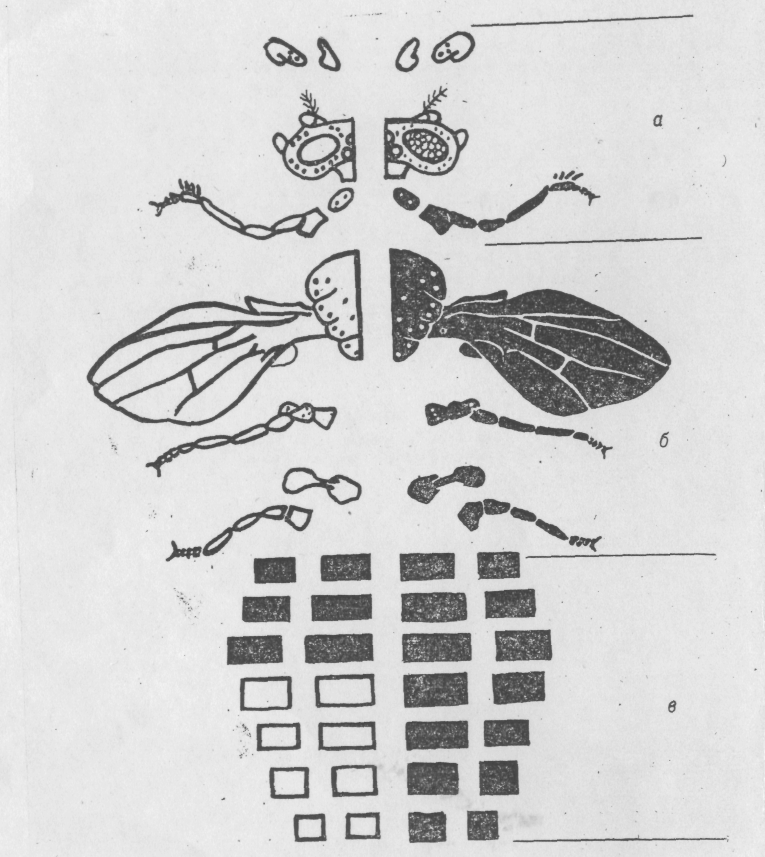
Рис. 6.4. Схематичне зображення зовнішніх частин тіла дрозофіли, що утворюються незалежно з окремих імагінальних дисків (а -голова; б -груди; в — черевце).
Для маркування «чоловічих» і «жіночих» частин тіла зазвичай використовують рецесивні анатомічні маркери (жовтий колір тіла, білі очі, змінені щетинки, тощо). Маркери не проявляються в тих частинах тіла, які мають генотип XX. У цьому випадку вони прикриті нормальними алелями. Однак вони видні в «чоловічих» частинах тіла, що втратили кільцеву X-хромосому, та мають одну X-хромосому. Завдяки цьому, відбираючи різних мозаїків, можна знайти, які частини тіла повинні бути мутантними, щоб сформувався певний мутантний поведінковий фенотип. Іншими словами, можна встановити, яка саме анатомічна структура або клітинна асоціація повинна бути порушена мутацією, щоб виявився ефект мутації на рівні цілісного організму.
Розв'язна здатність цього методу залежить від числа досліджуваних особин: чим більше це число, тим більше ймовірність, що серед них будуть мозаїки по самих близьких крапках.
Дослідження С. Бензера. Що стосується генетики поведінки, систематичне вивчення мозаїків D. melanogaster почалося з ініціативи С. Бензера. У 1970 р. ХХ століття він разом з І. Хота опублікував статтю за назвою «Генетичне розсічення нервової системи дрозофіли за допомогою мозаїків». У ній описано кілька способів одержання мозаїків, з них найбільш ефективний метод втрати в онтогенезі дрозофіли кільцевої Х-хромосоми.
У якості вихідного матеріалу брали гетерозиготних особин на стадії зиготи, у яких одна Х-хромосома кільцева — In(l)wvc, а інша, аналізована, несе обрану поведінкову мутацію й низку рецесивних маркерів В w sn f. Можна використовувати й інші маркери, що зачіпають покриви, пігментацію тіла й очей, форму щетинок.
Імовірність розриву й втрати кільцевої Х-хромосоми In(l)wvc у процесі клітинних поділів дуже велика й становить від 10% до 50%. У результаті цього й утворюються мозаїчні особини, які містять вихідні клітини - дипло-Х - і виниклі в результаті втрати кільцевої Х-хромосоми клітини гапло-Х з єдиною Х-хромосомою (аналізованою).
Чим раніше в онтогенезі відбудеться розрив і втрата Х-хромосоми In(l)wrc, тим більша кількість клітин тіла мухи буде гапло-Х. Якщо ця подія відбудеться вже при першому дробленні зиготи, то утворюється гінандроморф, у якого одна половина тіла буде жіночою, із клітинами дипло-Х, а інша — чоловічою, із клітинами гапло-Х. Ділянки тіла із клітинами гапло-Х легко впізнаються за рецесивними маркерами, що проявляються у гемізиготному стані.
Дроблення в дрозофіли відбувається за синцитіальним типом, і приблизно після 12 поділів ядра мігрують до поверхні яйця, утворюючи бластодерму. На стадії бластодерми окремі її ділянки вже детерміновані. З них формуються імагінальні диски тіла личинки, які, у свою чергу, дають початок цілком певним частинам тіла імаго. Окремі ділянки поверхні тіла (екзоскелета) і прилеглі до них тканини з високою ймовірністю складаються із клітин того самого клону. Наприклад, якщо ліва частина переднього грудного відділу мухи має кутикулу жовтого кольору, тобто складається із клітин гапло-Х, є всі підстави вважати, що відповідний відділ центральної нервової системи — ліва частина грудного синганглію — також буде з високою ймовірністю утворена клітинами гапло-Х. Це дозволяє знаходити так звані фокуси — структури, що зачіпаються аналізованими мутаціями.
У роботі, присвяченій мутаціям, що спричиняють дефекти зору, Хотта й Бензер прийшли до важливого висновку щодо автономності зорової системи кожного ока. При знятті ЕРГ у білатеральних гінандроморфів рецепторний потенціал нормального ока залишався нормальним, а рецепторний потенціал мутантного ока був аномальним. Удалося відібрати таких мозаїків, усе тіло яких було нормальним і тільки одне око було мутантним, тобто складалося із клітин гапло-Х з аналізованою мутацією в Х-хромосомі. ЕРГ, що знімалася з такого ока, незмінно виявлялася дефектною, причому характерною для даної мутації.
К. Ікеда й В. Каплан, користуючись аналогічною методикою, досліджували прояв шейкер-мутації Hklp в 77 мозаїчних особин. При цьому за допомогою мікроелектродної техніки характеризували електричну активність окремих мотонейронів. Ними переконливо доведена автономність окремих нейронів торакального синганглію. Без одержання мозаїчних особин подібного принципового результату навряд чи вдалося б досягти.
Хотта й Бензер у різних експериментах аналізували початкові етапи залицяння (переслідування самиць і вібрацію крил) і кінцеві етапи — спробу парування й саме спарювання. Усього ними вивчене в першій частині роботи 477 і в другій 208 мозаїчних особин із різними комбінаціями клітин дипло-Х і гапло-Х.
У першій частині роботи за допомогою бінокулярної лупи спостереження вели над окремими парами - самців гінандроморфів і самиць дикого типу C-S, - яких поміщали під годинникове скло. Результати досліджень показали, що контроль над орієнтацією мозаїків відносно самиць і вібрацією крил як елемента залицяння здійснюється церебральним ганглієм. Мозаїчні особини, у яких усе тіло, за винятком голови, було жіночим поводилися як самці — переслідували самиць і вібрували крилами.
У другій частині робіт спостереження за спарюваннями проводили за допомогою прилада, що включає одразу 10 камер. Учені виявили, що спроби до спарювання роблять гінандроморфи, в яких і голова, і торакс — самцеві. Для самого акту спарювання потрібна до того ж нормальна чоловіча будова геніталій.
У додатковій серії експериментів до мозаїчних особин підсаджували самців дикого типу. Наявність у мозаїків жіночих черевця й геніталій провокувало залицяння самців. Однак щоб мозаїчна особина поводилася як самиця, її центральна нервова система повинна була залишатися жіночою.
Використовуючи мозаїків, С. Бензер досліджував найрізноманітніші мутації, наприклад серію нефототаксисних мутацій, які належали до п'яти різних цистронів X-хромосоми. У нормі мухи характеризуються позитивним фототаксисом і рухаються в напрямку до світла прямо, при цьому світло рівної інтенсивності падає на обидва ока. Мутантні мухи з негативним фототаксисом воліють рухатися не до світла, а від нього. Нефототаксисні ж мутанти не проявляють ані позитивного, ані негативного фототаксису.
Цікавою є поведінка білатеральних мозаїків у яких одне око дефектне, тобто мутантне, а інше — нормальне. Вони реагують на світло, але на відміну від нормальних мух летять у напрямку до джерела світла не прямо, а по спіралі, постійно повертаючи своє мутантне око до світла, щоб інтенсивність світла, що падає на обидва ока, була однаковою (Рис. 6.5.).
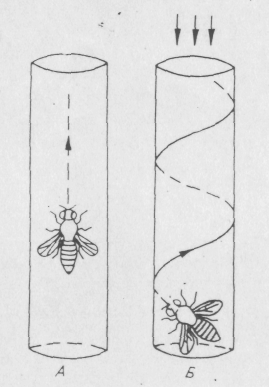
Рис. 6.5. Рух до джерела світла білатеральних мозаїків за фототаксисними мутаціями (А — нормальна муха; Б — білатеральний мозаїк з одним мутантним оком).
Дослідження електроретинограми мутантних за поведінкою мух підтвердило, що первинний фокус дії більшості нефототаксисних мутацій перебуває в самім оці. Усі мутації викликали різні відхилення в електроретинограмі, тому що ефект різних мутацій здійснювався різними шляхами. Причина одних відхилень — знижена світлочутливість фоторецепторних клітин, інші мутації викликали дегенерацію зорових паличок, у третіх — відхилення в електроретинограмі обумовлювалися дефектом передачі нервового сигналу від рецепторних клітин наступним нейронам.