

vse_metody_MIKROB
.pdfIII этап исследования: учет цитопатогенного действия (ЦПД) исследуемого вируса на культуру клеток Нер-2.
Под малым увеличением микроскопа отмечают дегенерацию клеток, свидетельствующую о размножении вирусов. Монослой разрушен, оставшиеся клетки округлые, без отростков, с темной зернистой цитоплазмой.
С целью идентификации вирусов производят постановку реакции нейтрализации (РН) в культуре клеток с аденовирусными диагностическими сыворотками к вирусам 3, 4 и 7 типов, т.к. они наиболее часто вызывают заболевания у людей.
Для постановки РН в три пробирки стерильной пипеткой вносят аденовирусные диагностические сыворотки 3, 4, 7 типов в объеме 0,2 мл. К каждой из этих сывороток добавляют другой стерильной пипеткой по 0,2 мл исследуемой вируссодержащей жидкости. Смеси оставляют для контакта на 1 час, затем каждой смесью в объеме 0,2 мл заражают флаконы с культурой клеток и добавляют по 0,8 мл среды 199.
Контроли реакции: контроль культуры клеток; контроль сывороток (культура клеток + смесь сывороток 3, 4, 7 типов); контроль вируса (культура клеток + вируссодержащая жидкость). Все флаконы (опытные и контрольные) помещают в термостат при температуре 370С на 48-72 часа.
IV этап исследования: учет РН в культуре клеток (таблица 6).
Таблица 6 – Схема учета РН в культуре клеток
Контроль |
Контроль |
Контроль |
Вирус + |
Вирус + |
Вирус + |
культуры |
смеси сы- |
вируса |
сыворотка |
сыворотка |
сыворотка |
клеток |
вороток |
|
3 типа |
4 типа |
7 типа |
|
|
|
|
|
|
Вконтроле культуры клеток и контроле смеси сывороток – монослой из нормальных клеток, т.е. отсутствует дегенерация.
Вконтроле вируса – дегенерация клеток за счет цитопатогенного действия исследуемого вируса.
Вопытных флаконах монослой из нормальных клеток остается в том флаконе, где произошла нейтрализация вируса, т.е. тип сыворотки совпал с типом инфицирующего агента.
Заключение о типе вируса сделайте самостоятельно.
9
Занятие №15
Тема. Бактериофаги. Генетика и изменчивость бактерий. Антибиотики.
Цель занятия. Изучить свойства бактериофагов, их практическое применение,
генетику и изменчивость бактерий, характеристику и спектр действия антибио-
тиков. Освоить методы определения чувствительности бактерий к антибиоти-
кам.
I.Теоретические знания:
1.Морфология и структура бактериофагов, их классификация.
2.Вирулентные и умеренные бактериофаги. Фазы взаимодействия вирулентно-
го бактериофага с клеткой. Практическое применение бактериофагов.
3.Организация генетического материала бактериальной клетки. Факторы вне-
хромосомной наследственности.
4.Виды генетической изменчивости. Использование генной инженерии в меди-
цине.
5.Антибиотики. Классификация антибиотиков по механизму антимикробного действия.
6.Методы определения чувствительности бактерий к антибиотикам. Осложне-
ния и последствия антибиотикотерапии.
II.Практические навыки:
1.Определение фаготипа бактерий.
2.Определение чувствительности бактерий к антибиотикам методом серийных разведений.
3.Определение чувствительности бактерий к антибиотикам диско-
диффузионным методом.
1

МЕТОДИЧЕСКИЕ УКАЗАНИЯ К ЗАНЯТИЮ
1. Морфология и структура бактериофагов, их классификация.
Бактериофаги (от греч. bacterion – бактерии, phagos – пожирающий) –
вирусы бактерий, вызывающие их гибель.
Бактериофаги были открыты в 1917 г. канадским ученым Ф. Д'Эррелем. Исследователь выделил из испражнений больных дизентерией фильтрую-
щийся агент, способный разрушать, лизировать дизентерийные бактерии. Последующие наблю-
дения показали, что бактериофаги встречаются повсеместно, где есть бактерии: в почве, сточных Ф. Д'Эррель водах, кишечном тракте человека и животных, гнойном отделяемом и других субстратах. По-
этому в широком смысле слова их часто называ-
ют просто фагами.
В зависимости от формы и структурной организации фаги подразделяют на пять морфологических групп (классификация А.С. Тихоненко) (рисунок 6):
I. Фаги I типа – нитевидной формы.
II. Фаги II типа – имеют головку и рудимент отростка.
III. Фаги III типа – имеют головку с коротким отростком.
IV. Фаги IV типа – имеют головку и длинный несокращающийся отросток.
V.Фаги V типа – имеют головку и длинный сокращающийся отросток.
Рисунок 6 – Морфологическая классификация бактериофагов по А.С.Тихоненко
2
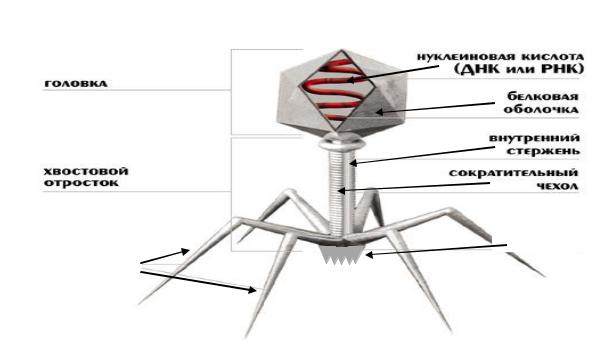
Самое сложное строение у фагов V группы.
Структура сложноустроенного фага (рисунок 7):
-головка, в которой содержится нуклеиновая кислота;
-воротничковая часть;
-отросток, сверху покрытый сократительным чехлом. На конце отростка находятся базальная пластинка и нити прикрепления для адсорбции фага на бактериальной клетке.
Оболочечные структуры фага имеют белковую природу.
воротничковая часть 
базальная
пластинка
нити
прикрепления
Рисунок 7 – Структура сложноустроенного фага
2.Вирулентные и умеренные бактериофаги. Фазы взаимодействия виру-
лентного бактериофага с клеткой. Практическое применение бактерио-
фагов.
В зависимости от характера взаимодействия с бактериальной клеткой, раз-
личают вирулентные и умеренные бактериофаги.
Вирулентные фаги способны вызывать взрывную продуктивную инфек-
цию. Проникнув в бактериальные клетки, они размножаются и вызывают лизис бактерий. Умеренные фаги чаще вызывают интегративную вирусную инфек-
цию, которая может переходить в продуктивную.
3
Фазы взаимодействия сложноустроенного вирулентного бактериофага
с клеткой:
 Адсорбция (отростковой частью фага) на клеточной стенке бактерий. В эту фазу рецепторы базальной пластинки и нитей прикрепления специфически взаимодействуют с определенными рецепторами клеточной стенки бактерий.
Адсорбция (отростковой частью фага) на клеточной стенке бактерий. В эту фазу рецепторы базальной пластинки и нитей прикрепления специфически взаимодействуют с определенными рецепторами клеточной стенки бактерий.
На бактериях, лишенных клеточной стенки (L-формы, микоплазмы), фаги не адсорбируются.
 Проникновение нуклеиновой кислоты фага в клетку: происходит сокра-
Проникновение нуклеиновой кислоты фага в клетку: происходит сокра-
щение чехла отростка и растворение с помощью фагового лизоцима неболь-
шого участка клеточной стенки бактерии. Затем ДНК из головки бактериофа-
га через канал отростка инъецируется (впрыскивается) в цитоплазму клетки,
при этом оболочка фага остается на поверхности бактериальной клетки.
 Синтез фаговых частиц (подобно синтезу вирусов в эукариотической клет-
Синтез фаговых частиц (подобно синтезу вирусов в эукариотической клет-
ке): происходит репликация нуклеиновой кислоты бактериофага с образова-
нием множества копий, а на рибосомах бактериальной клетки – синтез фаго-
вых белков головки и отростка.
 Композиция фаговых частиц: происходит сборка белковых оболочек и нуклеиновых кислот и формируются зрелые бактериофаги.
Композиция фаговых частиц: происходит сборка белковых оболочек и нуклеиновых кислот и формируются зрелые бактериофаги.
 Выход фагов из бактериальной клетки путем лизиса клетки изнутри. Он осуществляется за счет свободного лизоцима, выделяемого множеством фа-
Выход фагов из бактериальной клетки путем лизиса клетки изнутри. Он осуществляется за счет свободного лизоцима, выделяемого множеством фа-
гов, что приводит к гибели бактерий в результате ее осмотического лизиса.
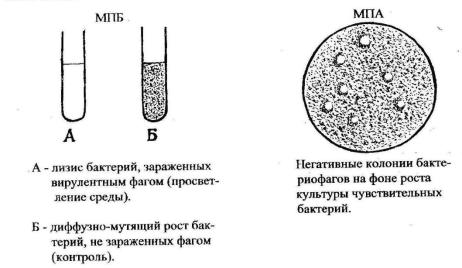
Репродукция вирулентного фага в популяции бактерий, выращенных в жидкой питательной среде (МПБ), сопровождается их лизисом и просветлени-
ем среды (рисунок 8а). В популяции чувствительных бактерий, выращенных сплошным газоном на плотной питательной среде (МПА), фаги образуют зоны очагового лизиса (рисунок 8б), которые называются «негативными колониями» или стерильными бляшками.
4
а) |
б) |
Рисунок 8 – Результат взаимодействия вирулентного бактериофага с бак-
териальной популяцией на жидкой и плотной питательной среде
Умеренные фаги чаще взаимодействуют с клеткой по типу интегративной вирусной инфекции: ДНК фага интегрируется с ДНК клетки и называется про-
фагом. Став частью хромосомы бактерии, профаг, при ее размножении, пере-
дается бактериальному потомству. Клетка, несущая профаг, называется лизо-
генной. Под влиянием различных факторов (УФ-света, некоторых химических веществ) связь профага с ДНК бактериальной клетки нарушается и профаг пе-
реходит в цитоплазму клетки, где размножается и ведет себя как вирулентный.
Практическое применение бактериофагов
I. Для диагностики инфекционных заболеваний используют метод фа-
готипирования, когда с помощью известного набора фагов определяют фаго-
вариант исследуемых бактерий. Метод основан на специфичности фагов, т.е.
способности взаимодействовать только с бактериями, имеющими специфиче-
ские рецепторы для адсорбции фага и лизиса этих бактерий. Фаготипирование используют для диагностики брюшного тифа, дизентерии, холеры, стафилокок-
ковых инфекций (рисунок 9).
5
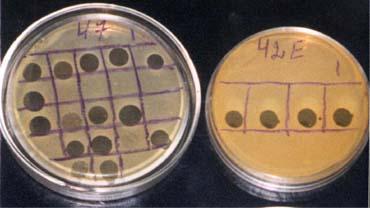
Рисунок 9 – Фаготипирование стафилококков с помощью набора различ-
ных типоспецифических фагов.
Метод фаготипирования имеет важное эпидемиологическое значение, т.к.
позволяет установить связи между источником инфекции и отдельными слу-
чаями заболевания. Выделение бактерий одного фаговарианта от разных боль-
ных указывает на общий источник их заражения.
II. Для лечения:
-стафилококковый бактериофаг (при гнойно-воспалительных заболева-
ниях, вызванных стафилококками);
-бактериофаг P.aeruginosa (при гнойно-воспалительных заболеваниях,
вызванных синегнойной палочкой);
-клебсиеллезный бактериофаг (при заболеваниях, вызванных клебсиел-
лами).
Комбинированные многокомпонентные препараты бактериофагов:
-коли-протейный бактериофаг (для лечения эшерихизов и дисбактерио-
зов, вызванных бактериями рода Proteus);
-пиобактериофаг (для лечения стафилококковой, стрептококковой, клеб-
сиеллезной, протейной, синегнойной инфекции и эшерихиозов);
-интести-бактериофаг (для лечения бактериальной дизентерии, сальмо-
неллезов, эшерихиозов, а также протейной, стафилококковой, энтеро-
кокковой и синегнойной инфекций).
6
Бактериофаги применяют местно путем аппликации на раневую или ожо-
говую поверхность, введением в полости (брюшную, плевральную, мочевой пу-
зырь), через рот, а также ректально. Соответственно способу применения пре-
параты бактериофагов выпускают в различных лекарственных формах (жидкая форма, таблетки, мази, свечи, аэрозоли). Перед назначением бактериофага
необходимо поставить пробу на чувствительность к нему выделенной
культуры микроорганизмов.
III.Для профилактики брюшного тифа и дизентерии у людей, контак-
тировавших с больным, используют брюшнотифозный и поливалентный дизен-
терийный бактериофаги.
3. Организация генетического материала бактериальной клетки. Факторы
внехромосомной наследственности.
Генетический материал бактериальной клетки представлен хромосом-
ной ДНК с гаплоидным набором генов. Хромосомная ДНК находится в супер-
спирализованной форме в виде кольца.
В бактериях могут присутствовать внехромосомные молекулы ДНК: плаз-
миды, транспозоны, вставочные последовательности.
Хромосомный и внехромосомный генетический материал свободно распо-
лагается в цитоплазме.
Факторы внехромосомной наследственности
Факторы внехромосомной наследственности (плазмиды, транспозоны,
вставочные последовательности) состоят из молекул ДНК, не являются жиз-
ненно важными для бактерий, но придают им новые свойства.
Плазмиды – кольцевидные молекулы ДНК, способные к саморепликации и несущие от 40 до 50 генов. Они находятся в автономном состоянии в цито-
плазме бактерий и способны к самопереносу из одной клетки в другую при конъюгации. Плазмиды кодируют свойства, дающие бактерии преимущества при попадании в неблагоприятные условия существования.
7
Плазмиды подразделяют на различные категории в зависимости от свойств, которые они кодируют у бактерий (таблица 7):
Таблица 7 – Классификация плазмид
Категории плазмид |
Свойства |
F-плазмида (от англ. fertility – |
Кодирует способность к переносу плазмид- |
плодовитость). |
ных и хромосомных генов при конъюгации |
|
бактерий через половые пили. |
R-плазмида (от англ. resis- |
Кодирует устойчивость бактерий к несколь- |
tance – противодействие) – |
ким антибиотикам и другим лекарственным |
плазмида резистентности. |
веществам. |
Плазмиды бактериоциноген- |
Кодируют синтез бактериоцинов (особых |
ности. |
белков), вызывающих гибель близкородст- |
|
венных бактерий. Например, бактериоцины |
|
E.coli вызывают гибель патогенных энтеро- |
|
бактерий. Бактериоцины могут быть полезны |
|
при лечении инфекций, вызванных антибио- |
|
тикорезистентными бактериями. |
Плазмиды патогенности. |
Кодируют синтез поверхностных бактериаль- |
|
ных антигенов, энтеротоксинов, гемолизинов |
|
и других факторов патогенности. |
Подвижные генетические элементы
Транспозоны – подвижные участки ДНК, которые способны перемещать-
ся внутри бактериальной хромосомы или между ДНК бактерий, плазмид и бак-
териофагов («прыгающие гены»). Помимо генов, кодирующих их перемещение по ДНК, транспозоны могут содержать структурные гены, обеспечивающие бактерию новыми свойствами (устойчивость к антибиотикам, токсинопродук-
ция и др).
Вставочные последовательности или инсерционные элементы (от англ. insertion sequens – вставочные последовательности) – простейший тип генети-
ческих элементов, мигрирующих между хромосомами, внутри хромосомы, ме-
жду хромосомой и плазмидами. Содержат гены, необходимые для их переме-
щения, новых свойств не кодируют.
8
Подвижные генетические элементы могут:
-распространять новые гены в популяции бактерий;
-координировать взаимодействие плазмид с хромосомой;
-вызывать изменения генов (мутацию, инактивацию) в местах их внедре-
ния в генетический материал.
Таким образом, внехромосомные молекулы ДНК бактериальной клетки способствуют разнообразным изменениям бактериального генома, появлению новых свойств и эволюционным изменениям микробной популяции в целом.
4. Виды генетической изменчивости. Использование генной инженерии в
медицине.
Мутации – это изменения последовательности нуклеотидов в бактериаль-
ной ДНК, которые ведут к изменению или утрате одного или нескольких фено-
типических признаков. Мутации у бактерий происходят с высокой частотой под влиянием очень многих внешних факторов, которые называются мутагена-
ми. Самые частые мутагены: УФ-, γ-лучи (физические), органические и неорга-
нические вещества (химические), транспозоны (биологические). Мутации бы-
вают: точечными (затрагивают один ген) и более протяженными (целый уча-
сток хромосомы). Они могут быть самопроизвольными и вызванными искусст-
венно. Мутации, которые приводят к возникновению новых полезных свойств,
обеспечивают выживаемость бактериальных популяции.
Генетические рекомбинации – это передача новых генов от одной клетки к другой с приобретением новых свойств, закодированных в этих генах. Гене-
тические рекомбинации отличаются друг от друга способом попадания в клетку новых генов. Виды генетических рекомбинаций: трансформация, конъюгация,
трансдукция, фаговая конверсия.
Трансформация у бактерий – форма генетической изменчивости, при ко-
торой бактерия-реципиент поглощает из внешней среды трофическим путем фрагменты ДНК погибшей бактерии-донора. Это приводит к образованию ре-
9