

Лекция 7
Фенотип и генотип. Взаимодействие аллельных и неаллельных генов в детерминации признаков. Количественная и качественная специфика проявления генов в признаках.
Термины «Фенотип» и «Генотип» ввел в 1909 году Вильгельм Иогансен. Он же сформулировал важные различия между этими понятиями. При этом утвердилось, что фенотип - это весь комплекс внешних и внутренних признаков организма, таких как форма, размеры, окраска, микроскопические и макроскопические особенности, химический состав, биохимические процессы.
Генотип - это совокупность всех генов организма с их аллельными формами. Однако следует иметь ввиду, что генотип - это не механическая сумма генов. В организме одновременно функционирует большое число генов из различных аллелей, в том числе расположенных в разных парах хромосом. В цепи реализации генов в признаки многие из них взаимодействуют друг с другом, что сказывается на проявлении каждого гена в признак. В этой связи генотип следует рассматривать как систему генов организма, определенным образом взаимодействующих между собой. Все механизмы взаимодействия генов в организме еще до настоящего времени не раскрыты. Однако уже можно говорить о том, что гены взаимодействуют друг с другом как непосредственно на уровне генетического материала, так и на уровне продуктов их функциональной активности. Во взаимодействии генов выделяют взаимодействие аллельных и неаллельных генов.
Взаимодействие аллельных генов
Аллельные гены находятся в одинаковых локусах гомологичных хромосом и отвечают за развитие одного и того же признака. В тоже время они являются альтернативными. Если в фенотипе проявляется один аллельный ген, то другой не проявляется. Взаимодействие аллельных генов происходит по типу:
1) полного доминирования;
2) неполного доминирования;
3) кодоминирования;
4) сверхдоминирования;
5) аллельного исключения;
6) межаллельной комплементации.
Полное доминирование. При полном доминировании имеет место проявление в фенотипе только одного гена из аллельной пары и проявление его не зависит от другого аллеля. Такой аллельный ген называют - доминантным. Он полностью подавляет действие другого аллеля и у гетерозигот при этом проявляется доминантный признак. Аллель, действие которого подавляется называется рецессивным. Рецессивный аллель проявляется в признак только в отсутствии альтернативного аллеля.
Примеров доминантных и рецессивных аллелей много у человека. Как доминантные, так и рецессивные гены могут нести патологический признак. Например, карий цвет глаз - признак доминантного гена, голубой - рецессивного; курчавые волосы - признак доминантного гена, прямые волосы - рецессивного; близорукость - признак доминантного гена, нормальное зрение - рецессивного; гемофилия - признак рецессивного гена.
Однако свойство доминантности и рецессивности аллелей (признаков) может носить относительный характер. В зависимости от условий его функционирования аллелельный ген может вести себя как доминантный, так и рецессивный. Например, отсутствие волос на голове (облысение) проявляет свойства рецессивного признака у женщин и доминантного у мужчин. То, что у женщин один из побочных эффектов применения в терапевтических целях мужского полового гормона тестостерона заключается в потере волос, указывает на участие мужского гормона в фенотипическом проявлении этого гена.
Неполное доминирование (промежуточное проявление). При гетерозиготности (Аа), когда в диплоидном наборе содержатся разные аллели, взаимодействие аллелей А и а не во всех случаях укладывается в рамки понятий о доминантности и рецессивности. Имеет место и промежуточное проявление, когда организм по признаку занимает среднее положение между доминантным и рецессивным признаком, среднее положение между обоими родителями. Примером неполного доминирования может быть наследование окраски цветков у ночной красавицы. При скрещивании растения с красным цветком (АА) и растения с белым цветком (аа), окраска гетерозиготы (Аа) имеет промежуточный характер - она розовая (см. схему).
P |
AA |
x |
aa |
||||
|
красн. |
|
бел. |
||||
G |
|
A |
|
|
|
a |
|
F1 |
|
Aa |
|
||||
|
|
розов. |
|
||||
При скрещивании гибридов F1 также проявляется промежуточный характер наследования признака у гетерозигот:
P |
Aa |
x |
Aa |
||||
|
розов. |
|
розов. |
||||
G |
A |
|
a |
|
A |
|
a |
F2 |
AA |
Aa Aa |
aa |
||||
|
1 |
2 |
1 |
||||
|
красн. |
розов. |
бел. |
||||
Механизм такого наследования сводится к ослаблению действия доминантного аллеля в присутствии рецессивного.
Кодоминирование. Аллельные гены могут в равной мере проявлять себя в фенотипе. Наличие в генотипе двух аллелей (у гетерозигот) ведет к возникновению нового выражения признака. Аллели, которые одинаково функционируют при их совместном присутствии в генотипе получили название кодоминантных, а само это явление называют кодоминирование. Кодоминирование аллелей у людей наблюдается в наследовании групп крови АВО. Аллели IA и IB кодоминантны, при взаимодействии их в генотипе формируется IV группа крови с генотипом IAIB. Такие гетерозиготы несут оба антигена - А и В.
Сверхдоминирование. При сверхдоминировании наблюдается более сильное проявление признака у гетерозиготных организмов по сравнению с исходными доминантными гомозиготными родительскими формами. Одним из объяснений сверхдоминирования является эффект гетерозиса, наблюдаемый у гибридов первого поколения. Гетерозис активно проявляется по количественным признакам, таким как плодовитость, продолжительность жизни, вес и др. Явление сверхдоминирования проявляется у гетерозигот по серповидно-клеточной анемии у человека. Гетерозиготы HbA/HbS обладают большей устойчивостью к тропической малярии по сравнению с доминантными гомозиготами HbA/HbА.
Аллельное исключение. При доминантности, рецессивности, кодоминировании, неполном доминировании результаты взаимодействия генов проявляются во всех соматических клетках организма. При аллельном исключении в части клеток организма, гетерозиготного по данному локусу, активен один аллель, тогда как в других клетках другой. Примером аллельного исключения может быть генетическая инактивация одной из Х-хромосом у особей гомогаметного пола (гиперхроматизирована в виде тельца Барра). Причем в разных клетках организма инактивируются разные Х-хромосомы. В связи с этим в одних клетках активен аллель, полученный с Х-хромосомой от матери, а в других клетках активен аллель, полученный с Х-хромосомой от отца. Известно, что в Х-хромосоме человека находится более тысячи генов, в том числе ген, контролирующий образование потовых желез. При наличии доминантного аллеля А это гена, в коже формируются нормальные потовые железы, а при наличии рецессивного аллеля а этого гена в Х-хромосоме в коже отмечается отсутствие потовых желез. Поскольку в различных клетках кожи может быть инактивирована либо одна, либо другая Х-хромосома, у женщин наблюдается мозаицизм в распределении потовых желез в коже в результате инактивации одной из Х-хромосом.
Межаллельная комплементация. Межаллельная комплементация относится к редко встречаемым формам взаимодействия аллельных генов. В этом случае возможно формирование нормального признака у организма, гетерозиготного по двум мутантным аллелям. Два разных мутантных аллеля совместно могут обеспечить нормальный фенотип. Такое взаимодействие проявляется чаще всего на уровне формирования четвертичной структуры белка, при объединении полипептидных цепей, образующих белок. Например, ген А отвечает за синтез нормального белка. Мутантный аллель а1 вызывает синтез измененного пептида, а мутантный аллель а2 определяет синтез другого, но тоже измененного пептида. Взаимодействие мутантных пептидов при формировании четвертичной структуры белка компенсирует измененные участки в них, что может привести к появлению белка с нормальными
свойствами (рис. 1).
|
|
|
||||||||||
Нормальный белок |
|
Нормальный белок |
Рис. 1. Межаллельная комплементация
При рассмотрении выше представленных форм взаимодействия аллельных генов учитывалось, что один и тот же ген представлен двумя аллелями А и а, В и в и так далее. Эти два состояния гена возникают при прямом и обратном мутировании. На самом деле один и тот же ген может изменяться в целый ряд состояний. При этом образуются многие варианты одного и того же гена. Например, ген А может мутировать: А→А1, А2, А3...Аn. Иногда таких аллельных форм может быть десятки.
Мутации одного и того же гена называют серией множественных аллелей, а само явление - множественный аллелизм. Надо иметь ввиду, что любой аллель серии может возникать мутантно непосредственно от аллеля дикого типа ( наиболее распространенный в природе аллельный ген) или любого другого аллеля данной серии. Любой аллель серии может мутировать как в прямом, так и обратном направлении.
Наследование генов серии множественных аллелей подчиняется менделевским закономерностям. При этом соблюдаются следующие положения:
1) серия множественных аллелей у каждого диплоидного организма может быть представлена одновременно только двумя любыми ее членами;
2) каждый из членов серии может полностью или неполностью доминировать над другими членами;
3) члены одной и той же серии действуют на один и тот же признак, одновременно они могут иметь множественный эффект.
Множественный аллелизм характеризует разнообразие генофондов вида и имеет большое значение в эволюции. Серия множественных аллелей обнаружена у различных растений и животных. Хорошо изучены серии аллелей генов формы цветка львиного зева, остистости колоса у пшеницы, окраски глаз у дрозофилы, окраски меха у кролика.

Рис. 2. Четыре фенотипа, возникающие при различных комбинациях аллелей гена окраски меха кролика
Множественный аллелизм можно проследить на примере кроликов по четырем генам окраски шерсти (рис. 2). Гены, отвечающие за окраску шерсти, у кроликов аллельны и находятся в гомологичных хромосомах в одних и тех же локусах. Ген дикой окраски-агути (С) доминирует над всеми остальными генами окраски. Ген шиншиловой (Сch) и гималайской (Ch) окраски доминируют над геном альбинизма (Ca). Гетерозиготы от шиншиловой и гималайской окраски дают светлую окраску.
Серии множественных аллелей обнаружены и у человека. В наследовании групп крови ABO участвует серия множественных аллелей, которая состоит из трех членов. Существует четыре группы крови системы ABO: O, A, B, AB. Они определяются тремя аллелями одного гена: IA, IB, iO. Аллели IA и IB доминантны по отношению к аллелю iO, но кодоминантны по отношению друг к другу. При наличии трех аллелей возможно шесть генотипов, но рецессивность iO сводит число групп крови к четырем:
Аллель |
|
Генотип |
|
Фенотип (группа крови) |
iO |
|
iOiO |
|
O (I) |
IA |
|
IAIA, IAiO |
|
A (II) |
IB |
|
IBIB, IBiO |
|
B (III) |
IA и IB |
|
IAIB |
|
AB (IV) |
Знание генетического предопределения групп крови имеет большое практическое значение, поскольку позволяет определить вероятность наследования той или иной группы крови в зависимости от генотипа родителей. Кроме того, позволяет раскрыть генетические механизмы несовместимости крови различных людей.
Взаимодействие неаллельных генов
Взаимодействие неаллельных генов разнообразно, сложно и по механизму до конца еще не раскрыто. Взаимодействие неаллельных генов может происходить по типу:
1) комплементарности;
2) эпистаза;
3) полимерии.
Комплементарность. При комплементарном взаимодействии генов признак развивается при наличии в генотипе двух доминантных неаллельных генов, каждый из которых не имеет самостоятельного проявления по этому признаку. К комплементарным генам относятся такие гены, которые при совместном присутствии в генотипе в гомо- или в гетерозиготном состоянии (A_B_) дают развитие нового признака. Действие же каждого гена в отдельности (AAbb) или (aaBB) данный признак не воспроизводит.
Примеров комплементарного взаимодействия доминантных неаллельных генов много у растений и животных. Наглядным примером комплементарного взаимодействия генов является наследование форм гребня у кур (рис. 3).
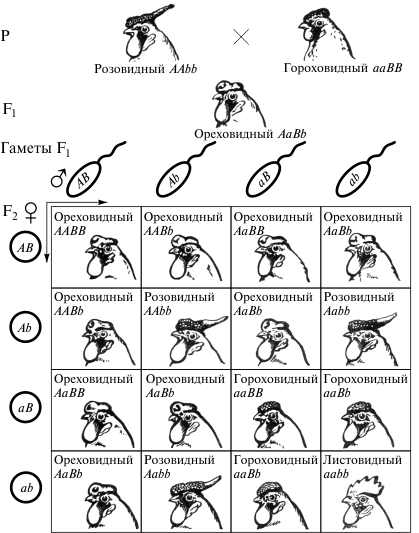
Рис. 3. Взаимодействие неаллельных генов при скрещивании кур с разными типами гребней
Две пары неаллельных генов в этом случае обеспечивают появление четырех форм гребня у кур: ореховидного, розовидного, гороховидного и листовидного. При скрещивании кур с гороховидным гребнем (AAbb) и розовидным гребнем (aaBB) все гибриды первого поколения имеют новую форму проявления гребня - ореховидную. Это новое проявление признака возникает благодаря взаимодействию двух доминантных неаллельных генов А и B в дигетерозиготах (AaBb). При скрещивании дигетерозигот во втором поколении происходит расщепление по фенотипу 9:3:3:1. Но ореховидным гребнем будут обладать только особи, в генотипе которых присутствует два доминантных неаллельных гена одновременно: AABB, AABb, AaBB, AaBb.
Классическим примером комплементарного взаимодействия генов является наследование трех типов окраски шерсти у мышей: дикой (серой) или «агути», черной и белой. Серые составляют большинство в природных популяциях. Такая окраска развивается вследствие взаимодействия имеющихся в генотипе двух доминантных неаллельных генов A и B. Один ген А контролирует синтез пигмента, другой B обеспечивает скопление пигмента в основании и конце волос. Их совместное присутствие в генотипе ведет к формированию признака серой окраски шерсти. Доминантные гомозиготы по этим генам будут иметь серую окраску шерсти. Гибриды F1 от них (AaBb) также будут иметь серую окраску вследствие взаимодействия доминантных аллелей обоих генов. Если скрестить гибридов F1 между собой, то 9/16 особей в F2 будут с генотипом A_B_, следовательно, с серой окраской шерсти (рис.4).
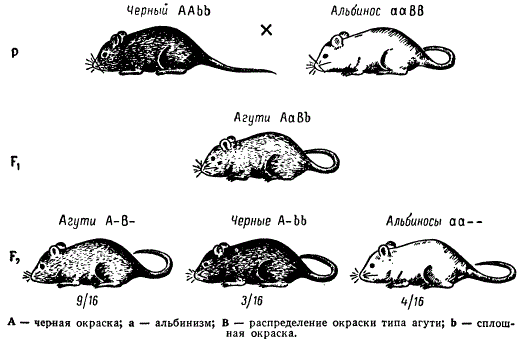

Рис. 4. Наследование трех типов окраски у мышей, как пример комплементарного взаимодействия неаллельных генов
Эпистатическое действие генов. Эпистаз - подавление действия одного гена другим не аллельным геном. Эпистатический ген (супрессор) действует на подавляемый гипостатический ген по принципу близкому к доминантности - рецессивности. Разница состоит в том, что эпистатический и гипостатический гены не являются аллельными, то есть занимают различные локусы в гомологичных или негомологичных хромосомах. Эпистаз широко распространен в природе, в некоторых случаях изучены биохимические механизмы эпистатических взаимодействий. Гены супрессоры могут быть доминантными и рецессивными, поэтому эпистаз делят на два типа: доминантный и рецессивный. Под доминантным эпистазом понимают подавление доминантной аллелью одного гена фенотипического проявления аллельной пары другого гена (А подавляет действие B и b).
Примером доминантного эпистаза может служить наследование окраски плодов у некоторых тыкв. Этот признак контролируется геном, представленным двумя аллелями: доминантным - желтой окраски B и рецессивным - зеленой окраски b. Но если в генотипе растения присутствует доминантный аллель другого неаллельного гена А, то в присутствии любого из аллелей первого гена развивается белая окраска. Ген А выступает в роли гена-супрессора. При скрещивании растений дигетерозиготных по генам A и B, в отличие от обычного дигибридного скрещивания, наблюдается расщепление гибридов по признаку окраски плодов в отношении 12 (белые): 3 (желтые): 1 (зеленые). Это объясняется эпистатическим действием аллеля А (белая окраска) на аллель B (желтая окраска) и b (зеленая окраска).
Схема скрещивания при эпистатическом взаимодействии генов:
Р ♀ AaBb x ♂ AaBb
♂ ♀ |
AB |
Ab |
aB |
ab |
AB |
AABB бел. |
AABb бел. |
AaBB бел |
AaBb бел |
Ab |
AABb бел |
AAbb бел |
AaBb бел |
Aabb бел |
aB |
AaBB бел |
AaBb бел |
aaBB жёлт |
aaBb жёлт |
ab |
AaBb бел |
Aabb бел |
aaBb жёлт |
aabb зелён |
Под рецессивным эпистазом понимают такой тип взаимодействия, когда рецессивный аллель одного гена, будучи в гомозиготном состоянии (аа), не дает проявляться фенотипически доминантной или рецессивной аллели других генов. В качестве примера рецессивного эпистаза можно привести объяснение бомбейского феномена.
В семье женщины с первой группой крови (ioio) и мужчины со второй группой крови (IAIA) родился ребенок с IV группой крови (IAIB). С точки зрения взаимодействия аллельных генов групп крови - это невозможно. Исследование этого феномена показало, что для синтеза антигенов групп крови A и B в системе ABO еще необходим белок-предшественник, за синтез которого отвечает доминантный ген H. Если в генотипе рецессивная аллель этого гена в гомозиготном состоянии (hh) сочетается с доминантным аллелем гена групп крови IA или IB, антигены A и B не синтезируются и фенотипически такие люди имеют 1(O) группу крови ioio. При исследовании оказалось, что женщина унаследовала от матери ген IВ, а от отца ген io. Но проявил свое действие только ген io, поэтому считалось, что женщина имеет 1(O) группу крови. Ген IВ был подавлен рецессивным аллелем h, находящимся в гомозиготном состоянии. Подавленный ген IВ проявил свое действие во втором поколении и ребенок имел IV группу крови.
Полимерия. Полимерия - это явление, когда один и тот же признак определяется несколькими доминантными неаллельными генами. Такие гены получили название полимерных. Поскольку они влияют на один и тот же признак, их обозначают одной латинской буквой с указанием индекса для разных генов: A1; A2; A3 и т.д.
Полимерия - это явление полигенной наследственности. При этом два или более доминантных гена контролируют содружественно развитие одного признака. Чем больше доминантных полимерных генов в генотипе, тем сильнее проявляется признак. Полимерному наследованию подчинена передача по поколениям количественных признаков, поскольку при взаимодействии полимерных генов происходит суммирование влияния их на развитие признака. К количественным признакам относятся: масса тела, рост, возможно умственные способности, цвет кожи.
По полимерному типу у людей наследуется интенсивность окраски кожных покровов, зависящая от количества синтезируемого пигмента меланина в клетках кожи. Генетический контроль количества образуемого пигмента осуществляется независимо четырьмя полимерными генами. Синтез пигмента предопределен доминантными аллелями: P1, P2, P3, P4. Наличие в генотипе человека восьми доминантных аллелей (P1P1P2P2P3P3P4P4) обуславливает максимальную пигментацию кожных покровов, отличающую африканских негров. Гомозиготность по рецессивным аллелям четырех генов (p1p1p2p2p3p3p4p4) характеризуется минимальным уровнем пигментации, отличающим представителей европеоидной расы. При браке африканцев с европеоидами рождаются мулаты, которые будут тетрагетерозиготами (P1p1P2p2P3p3P4p4) по полигенам синтеза пигмента. По количеству образующегося пигмента они будут занимать промежуточное (среднее) положение по сравнению с родителями. При браках муллатов между собой, благодаря независимому комбинированию аллелей четырех полимерных генов, количество доминантных аллелей в генотипе детей может колебаться от 0 до 8. Следовательно дети могут рождаться от таких браков от максимально светлой до максимально темной пигментации кожи (рис. 5).
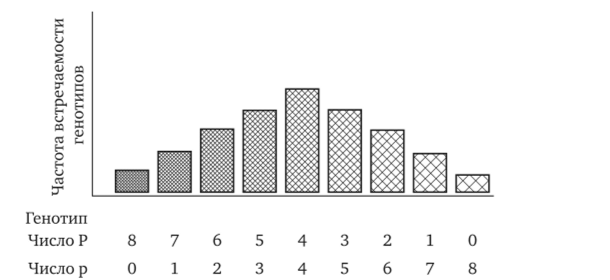
Рис. 5. Зависимость пигментации кожных покровов у человека от числа в генотипе доминантных аллелей
Количественная и качественная специфика проявления генов в признак
Проявление действия того или иного гена имеет свои характерные особенности. Один и тот же аллель у разных организмов может проявлять свой эффект различным образом. Это обусловлено состоянием генотипа в целом и условиями среды, в которых он существует. Фенотипическое проявление гена может варьировать по степени выраженности признака. Степень выраженности признака у разных особей, которые являются носителем соответствующего гена, получило название экспрессивность. Большинство организмов, носителей определенного аллеля, имеют среднюю степень развития признака.
Один и тот же признак может проявляться у одних организмов и отсутствует у других, имеющих тот же ген. Это явление было названо пенетрантностью. Таким образом, пенетрантность - это количественный показатель фенотипического проявления гена (степень проявления гена). Пенетрантность измеряется по проценту особей в популяции, у которых проявляется в признаке данный аллель. При полной пенетрантности (100%) ген проявляет свое действие у каждой особи, которая им обладает. При неполной пенетрантности (ниже 100%) ген проявляет свой фенотипический эффект не у всех особей. Многие гены имеют полную пенетрантность. Например, в опытах Г. Менделя все горошины, несущие доминантный аллель (определяющий желтую окраску как в гомозиготном, так и гетерозиготном состоянии), были желтыми, а все горошины гомозиготные по аллелю, определяющему зеленую окраску - зелеными.
Примером неполной пенетрантности может служить проявление доминантного гена, вызывающего хорею Гентингтона у человека. Это заболевание приводит к дегенеративным изменениям нервной системы. Люди, несущие этот доминантный ген, заболевают в различном возрасте, а у некоторых носителей он так и не проявляется. Они умирают от старости или других причин, не заболев хореей Гентингтона.
Причинами, вследствие которых ген проявляется у одних особей и не проявляется у других, а так же имеет варьирующую экспрессивность, могут быть особенности генотипов особей, влияние на эти гены других генов, различная реакция генотипа на факторы среды.
В этом и предыдущих разделах мы рассматривали, главным образом, взаимодействия между генами, влияющими на один признак. Но поле действия гена может быть гораздо шире, чем определение какого-то конкретного признака. Один ген может оказывать влияние на развитие нескольких признаков, проявлять множественное действие. Это явление получило название плейотропия. Например, Г. Мендель отмечал, что один из изучаемых им генов воздействовал одновременно на окраску цветов (белая или красная), окраску семян (серая или коричневая) и окраску пазух листьев (наличие или отсутствие пятен). У дрозофилы, гомозиготные по аллелю vestigial (ag) особи, имеют зачаточные крылья. Это основное фенотипическое проявления данного гена. Однако такие мухи отличаются от мух дикого типа и по многим другим признакам: у них изменены жужжальца, несколько изменены репродуктивные органы, снижена плодовитость и продолжительность жизни и т.д. Примером плейотропного гена у человека служит доминантный ген, определяющий синдром Марфана. У взрослых людей с синдромом Марфана отмечается высокий рост, за счет удлиненных конечностей с длинными тонкими (паукообразными) пальцами, врожденный подвывих хрусталика глаза, нередко врожденные пороки сердца.
Поскольку на уровне первичного действия гена каждый структурный ген имеет единственную функцию - кодирование одного полипептида, плейотропный эффект является, очевидно, отражением интегрированности процессов развития, в которых продукт одного биохимического пути может в конечном итоге оказывать влияние на множество путей развития.
Итак, проявление генов в признаки определяется, как правило, всем генотипом и каждый ген может действовать на развитие многих признаков. С этими механизмами связано явление генокопии. Сущность генокопии заключается в том, что ряд сходных по внешнему проявлению признаков, в том числе патологических, может вызываться различными неаллельными генами.
Таким образом, в этой лекции была представлена довольно сложная картина взаимодействия генов генотипа в детерминации признаков организма. Все это определяет развитие соответствующего фенотипа. Однако надо иметь ввиду, что в фенотипе никогда не реализуются все генетические возможности, фенотип каждого организма лишь частичный случай проявления его генотипа в конкретных условиях развития. Доказательством этого служит тот факт, что особи со сходным генотипом, например, однояйцевые близнецы, в разных условиях развития могут сильно различаться во взрослом состоянии. В формировании фенотипа конкретного человека играют большое значение и социальные условия, в которых он формируется. Одним словом, в формировании фенотипа заложен полифакториальный принцип. Сущность этого принципа в диалектическом единстве генетических и средовых факторов, определяющих фенотипические признаки.