

Лекция 15
Индивидуальное развитие организма (онтогенез). Периодизация онтогенеза. Предэмбриональный и эмбриональный периоды развития.
Индивидуальное развитие организма - это совокупность процессов, которые обеспечивают возникновение и динамику развития организма во времени. Индивидуальное развитие понятие широкое, которое в настоящее время сформировалось в отдельную область науки о живом – биологию развития, изучающую закономерности онтогенетического развития организма. Биология развития сформировалась путем синтеза ряда биологических дисциплин: эмбриологии, генетики, молекулярной биологии, цитологии, биохимии. В настоящее время биология развития изучает закономерности онтогенеза на молекулярном, генетическом, клеточном, тканевом, органном и организменном уровнях. На сегодняшний день наши знания все еще ограничены о закономерностях индивидуального развития. Например, не существует достаточно аргументированных теорий, которые могли бы объяснить каким образом закодирована и как реализуется информация об особенностях пространственного строения органов высших организмов. Какой таинственный механизм заставляет принять организм именно ту форму, которая свойственна данному виду, причем с поразительной точностью. Единственное, что не вызывает сомнения, это то, что основу процесса развития составляет наследственная информация, полученная потомками от родителей через половые клетки. И развитие в этой связи надо рассматривать как процесс реализации генетической информации.
Прежде чем давать характеристику индивидуального развития необходимо отметить, что развитие организмов может быть личиночное (непрямое) и прямое. Личиночное развитие характеризуется тем, что в развитии организма имеется одна или несколько личиночных стадий. Личинки ведут активный образ жизни, сами добывают пищу. У личинок имеется временные (провизорные) органы с помощью которых осуществляются жизненные функции личинок. У взрослого организма эти органы отсутствуют. В дальнейшем происходит метаморфоз – превращение личинки во взрослый организм. При прямом развитии из оплодотворенной яйцеклетки происходит постепенное развитие взрослого организма. При этом также имеют место на определенных этапах провизорные (зародышевые) органы, которые функционируют у зародыша и отсутствуют во взрослом состоянии. У млекопитающих и человека к ним относятся зародышевые оболочки (хорион, амнион, желточный мешок, аллантоис), а также плацента.
Индивидуальное развитие организма представляет непрерывный процесс развития. Однако в нем выделяют отдельные периоды и стадии в зависимости от смены преобладающих молекулярных, клеточных и системных механизмов, а также от характера взаимоотношений организма с внешней средой. Во-первых, выделяют три периода: предэмбриональный (предзародышевый); эмбриональный и постэмбриональный. Предэмбриональный период – это период развития половых клеток. Эмбриональный период – это период от оплодотворения яйцеклетки и образования зиготы до момента рождения, что касается млекопитающих и человека, или выхода из яйцевых и защитных оболочек, что касается других организмов. Постэмриональный период – это период с момента появления на свет до естественной гибели организма.
Предэмбриональный период. Это период развития половых клеток (гаметогенез). Этот процесс мы подробно рассматривали ранее. Тем не менее, следует назвать отдельные этапы развития половых клеток: 1) обособление первичных половых клеток от других (соматических) клеток организма; 2) размножение половых клеток, называемых на этой стадии гаметогониями; 3) рост половых клеток; 4) созревание половых клеток (мейоз); 5) формирование.
Необходимо отметить некоторые особенности развития яйцеклеток, которые связаны с обеспечением ранних этапов развития зародыша. Одной из таких особенностей овогенеза является интенсивное накопление огромного запаса компонентов аппарата трансляции (биосинтеза белка). У многих животных в ооците в короткие сроки накапливается огромное количество рибосом. Этому способствует умножение или амплификация генов рибосомных РНК яйцеклетки. При обычной репликации ДНК удваивается вся ДНК хромосомы, тогда как при амплификации копируются только избранные участки, в данном случае участки ДНК, где находятся гены рибосомных РНК. Копии ДНК отделяются от хромосомы, в результате формируются большое число дополнительных ядрышек, которые интенсивно продуцируют рибосомы. Кроме того, при овогенезе накапливаются тРНК, интенсивно синтезируются и депонируются иРНК. Таким образом, в яйцеклетке запасаются все компоненты, необходимые для синтеза белка, которые будут использоваться на ранних стадиях развития эмбриона. Вторая особенность овогенеза – образование и накопление в цитоплазме яйцеклетки питательных веществ (желток, углеводы, жиры, липиды, витамины, минеральные соли), которые используются также на ранних стадиях развития зародыша. Основную массу желтка составляют белки. В состав желтка, накапливаемого в овогенезе, входит примерно 90% всего белка клетки.
Для яйцеклетки характерна ооплазматическая сегрегация. Непосредственно перед оплодотворением начинаются и продолжаются после оплодотворения в зиготе интенсивные перемещения составных частей (компонентов) цитоплазмы яйцеклетки разного химического состава. Это явление получило название ооплазматическая сегрегация. Эти процессы создают определенную пространственную организацию будущего зародыша и определяют начальные этапы его дифференцировки, о чем будет говориться ниже.
Эмбриональный период. В эмбриональном периоде выделяют ряд отдельных стадий: 1) стадия оплодотворения; 2) стадия зиготы; 3) стадия дробления (образование однослойного зародыша); 4) стадия гаструляции (образование двух- и трехслойного зародыша); 5) стадия гисто-, органогенеза).
Стадия оплодотворения. Оплодотворение – это процесс слияния половых клеток. Выделяют наружное и внутреннее оплодотворение. У большинства животных, обитающих в воде, гаметы выделяются наружу и оплодотворение происходит вне тела матери. У животных, ведущих наземный образ жизни, семенная жидкость вводится в организм самки во время полового акта. Там и происходит оплодотворение. Однако процесс оплодотворения весьма сложный, включает ряд фаз и сопровождается выраженными изменениями в яйцеклетке и сперматозоиде. В процессе оплодотворения можно выделить три фазы: 1) сближение гамет; 2) активация гамет; 3) слияние гамет (сингамия). Сближение гамет сводится к дистантному взаимодействию между яйцеклеткой и сперматозоидом. Первоначально предполагали, что движение сперматозоида к яйцу носит целенаправленный характер и происходит благодаря хемотаксису. Предположение о хемотаксическом механизме сближения не подтвердилось , за исключением отдельных случаев, описанных у некоторых видов гидромедуз и высших споровых растений. На самом деле сближение сперматозоида с яйцом обеспечивается совокупностью ряда неспецифических факторов, повышающих вероятность их столкновения: 1) координирование процессов гаметогенеза у самца и самки и одновременность наступления стадии готовности к оплодотворению; 2) приспособления, связанные с оплодотворением и совокуплением, которые обеспечивают попадание созревших половых клеток в места, где происходит процесс оплодотворения; 3) избыточная продукция сперматозоидов по сравнению с числом женских половых клеток; 4) крупные размеры яйца, способствующие повышению вероятности столкновения со сперматозоидом. Не исключено, что существуют и другие факторы, однако и перечисленных достаточно, чтобы обеспечить оплодотворение всех созревших яиц животного. Кроме того, яйцеклетки и сперматозоиды вырабатывают химические вещества, которые возможно участвуют в обеспечении их сближения и взаимодействия. Эти вещества были названы гамоны.
Активация гамет начинается после соприкосновения сперматозоида и яйцеклетки. Процесс активации сперматозоида называется – акросомная реакция, яйцеклетки – кортикальная реакция. Принципы оплодотворения примерно одни и те же у разных видов животных. Но есть некоторые особенности у разных видов, которые, в частности, зависят от строения оболочек яйца. Нас конечно интересуют млекопитающие и человек. У млекопитающих и человека сперматозоид должен преодолеть несколько слоев фолликулярных клеток, блестящую оболочку и плазматическую мембрану яйцеклетки. Когда сперматозоид соприкасается с поверхностью яйца он активируется (акросомная реакция): в области акросомы сперматозоида разрушается плазматическая мембрана, из сперматозоида выходят гидролитические ферменты (сперматолизины), которые разрушают контакты между фолликулярными клетками, блестящую оболочку и сперматозоид достигает плазматической мембраны яйца. В месте контакта плазматическая мембрана яйца разрушается, плазматические мембраны сперматозоида и яйцеклетки сливаются и сперматозоид оказывается внутри яйцеклетки.
При соприкосновении сперматозоида и яйца активируется и яйцеклетка (кортикальная реакция). При кортикальной реакции повышается в яйце содержание ионов кальция, что сопровождается активированием биохимических процессов в клетке. Надо иметь в виду, что при оплодотворении сперматозоид прикрепляется в определенном месте блестящей оболочки, в том месте, где имеется специфический рецептор к сперматозоиду, который синтезируется яйцом. Только после фиксации сперматозоида на этом рецепторе он проникает через блестящую оболочку, плазматическую мембрану и попадает внутрь яйцеклетки, завершая процесс оплодотворения. Затем рецептор изменяется и тем самым создается препятствие для фиксации и проникновения внутрь других сперматозоидов. Это препятствует полиспермии у млекопитающих и человека. У ряда животных, в частности в яйцеклетках морского ежа, полиспермии препятствует изменение электрического потенциала плазматической мембраны яйца, а также формирование оболочки оплодотворения после проникновения одного сперматозоида.
Стадия зиготы. После проникновения сперматозоида в яйцеклетку начинается стадия зиготы. Заканчивается мейоз, формируется ядро яйцеклетки, которое нередко называют женским пронуклеусом. В период завершения женского мейоза ядро головки сперматозоида преобразуется. При этом оно увеличивается, происходит набухание и разрыхление хроматина, также как и в ядре яйцеклетки. Происходит перемещение пронуклеусов по цитоплазме. В процессе движения пронуклеусов и разрыхления хроматина в каждом из них реплицируется ДНК. Перед завершением сближения в каждом из пронуклеусов количество ДНК возрастает вдвое. Но общего ядра у млекопитающих и человека не образуется. Конечная стадия сближения пронуклеусов – это образование одной метафазной пластинки деления зиготы. В зиготе интесивно продолжается ооплазматическая сегрегация.
Стадия дробления. После объединения хромосомных наборов обоих пронуклеусов тотчас начинается ряд последовательных митотических делений зиготы, заканчивающихся формированием многоклеточного однослойного зародыша – бластулы. Клетки дробящегося яйца называют бластомерами. Побуждает зиготу делиться несоответствие величины ядерно-цитоплазматического отношения зиготы величинам, которые характерны для соматических клеток. За первым делением дробления следует серия следующих делений, так называемых делений дробления. Для всех этих делений характерны общие свойства: 1) разделившиеся клетки зародыша не растут. Суммарный объем и масса всех возникших клеток не превышает объема и массы яйцеклетки сразу после оплодотворения; 2) количество ДНК в ядрах удваивается после каждого деления, как при обычном митозе; 3) дробление зигот фактически у всех видов характеризуется асинхронностью, т. е. не одинаковыми циклами деления. У млекопитающих уже со второго деления, дробление идет асинхронно. Строение бластулы зависит от типа дробления яйца, а тип дробления во многом определяется количеством и распределением желтка в яйце, т.е. типом яйцеклетки.
Все типы дробления можно разделить на полное (голобластическое) и неполное (меробластическое). При полном дроблении дробится вся яйцеклетка, при неполном частичное дробление. Голобластическое (полное) дробление делят на: равномерное и неравномерное. При полном равномерном дроблении образуются бластомеры равной величины, при полном неравномерном - бластомеры оказываются разной величины. Полное равномерное дробление характерно для изолецитальных яиц, например, ланцетника. Полное неравномерное дробление характерно для умеренно телолецитальных яиц амфибий. Неполное (меробластическое) дробление также подразделяется на два: дискоидальное дробление резко телолецитальных яиц (птиц и некоторых рептилий) и поверхностное дробление центролецитальных яиц (членистоногих).
Рассмотрим на примере ланцетника полное равномерное дробление (рис. 1). Ему характерен радиальный тип дробления. Это значит, плоскости деления клеток в последовательных делениях проходят перпендикулярно друг другу. В результате бластомеры располагаются симметрично друг над другом. Первая борозда делит яйцо меридиально на два бластомера, а затем вторая борозда проходит тоже по меридиану, перпендикулярно первой борозде. Третья борозда проходит экваториально, образуется 8 бластомеров. Затем происходит чередование экватореальных и меридиальных борозд. На стадии 16 бластомеров образуется плотный комочек клеток. Зародыш на этой стадии называется морулой. В дальнейшем в результате дробления зародыш приобретает вид шара с полостью внутри. Стенка бластулы образована одним слоем клеток – бластодерма. Все бластомеры при этом имеют одинаковый размер. Полость бластулы – бластоцель.Такая бластула с тонкой однослойной стенкой и обширным бластоцелем получила название целобластула или типичная бластула.

Рис. 1. Полное равномерное дробление (у ланцетника)
Голобластическое неравномерное дробление характерно для умеренно телолецитальных яиц амфибий (рис. 2). Неравномерное распределение желтка и образование вследствие этого вегетативного и анимального полюсов ведет к задержке дробления у вегетативного полюса. Когда борозда дробления доходит до вегетативного полюса, ее дальнейшее продвижение замедляется настолько значительно, что последующие клеточные деления начинаются на анимальном полюсе раньше, чем на вегетативном успевает закончиться первое деление. В результате бластомеры имеют неодинаковую величину: на анимальном полюсе они мелкие (микромеры), на вегетативном полюсе крупные (макромеры), бластоцель при этом смещена в анимальное полушарие. Бластулу такого типа называют амфибластулой.
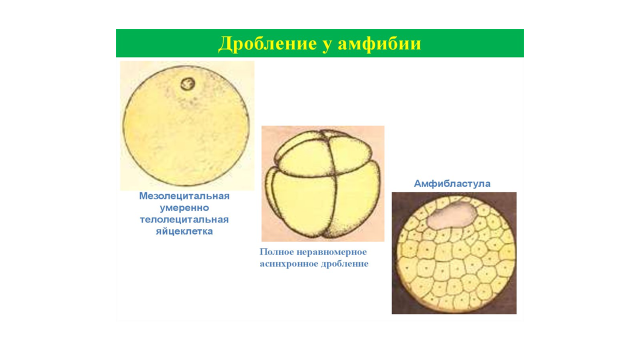
Рис. 2. Голобластическое неравномерное дробление (у амфибий)
При неполном дискоидальном дроблении резко телолецитальных яиц дробление идет только на анимальном полюсе. В результате образуется дискобластула, а бластоцель располагается под слоем бластодермы на анимальном полюсе в виде щели (рис 3).

Рис. 3. Неполное дискоидальное дробление (у птиц)
При неполном поверхностном дроблении центролецитальных яиц членистоногих ядра яиц многократно делятся и перемещаются к периферии, где в цитоплазме нет желтка. Образуется на периферии один слой бластомеров, а бластоцель заполнен желтком (рис. 4).

Рис. 4. Неполное поверхностное дробление (у членистоногих)
Дробление и формирование бластулы у млекопитающих. У высших (плацентарных) млекопитающих яйца изолецитальные (алецитальные) – имеется небольшое количество желтка. Дробление полное, неравномерное, почти с самого начала асинхронное. В результате дробления образуется морула, состоящая из 16-32 клеток. В моруле очень скоро выделяется слой светлых наружных клеток и более темная плотная масса внутренних клеток. Из наружного слоя впоследствии развивается особая внезародышевая ткань – трофобласт, а из внутренней массы (эмбриобласта или зародышевого узелка) формируется сам зародыш и его провизорные органы. В конце дробления в зародыше возникает обширная полость (бластоцель). На этой стадии зародыш млекопитающих называется бластоцистой. Наружный слой клеток бластоцисты представляет собой трофобласт, из которого позднее разовьется наружная оболочка плода – хорион. Трофобласт играет важную роль в имплантации зародыша в стенку матки. У анимального полюса бластоцисты располагается эмбриобласт (рис. 5).
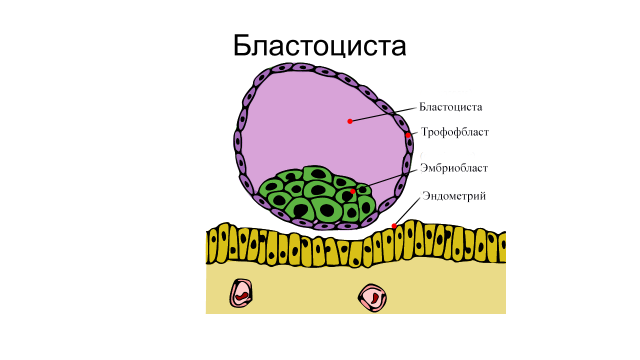
Рис. 5. Бластоциста млекопитающих
Стадия гаструляции. Одно из главных изменений, происходящих в начале гаструляции – это приобретение клетками способности к направленным морфогенетическим перемещениям. После прохождения стадии бластулы в зародыше начинается интенсивное перемещение как отдельных клеток, так и обширных участков стенки бластулы. Это приводит к тому, что более менее однородный перед этим зародыш расчленяется на два или три слоя, которые называются зародышевыми листками. У губок и кишечнополостных формируется два зародышевых листка: поверхностный листок- эктодерма и внутренний листок – энтодерма. У всех остальных животных формируется еще и третий, средний зародышевый листок – мезодерма, который располагается между двумя первыми. Способы гаструляции довольно разнообразны, отчасти они связаны со строением бластулы. Различают четыре основных способа гаструляции: инвагинация (впячивание), деламинация (расслоение), иммиграция (вселение), эпиболия (обрастание).
При инвагинации часть однослойной бластодермы (стенки бластулы) впячивается внутрь бластоцеля. В результате чего из однослойного полого шара образуется двуслойная чаша (двуслойный зародыш). Наружный слой клеток двуслойного зародыша представляет эктодерму, внутренний слой клеток – энтодерму. Энтодерма выстилает новую полость двуслойного зародыша, называемую - гастроцель или полость первичной кишки. Отверстие, ведущее в гастроцель принято называть бластопором или первичным ртом. Края бластопора называют губами бластопора. У первичноротых животных (черви, моллюски, членистоногие) бластопор в дальнейшем превращается в ротовое отверстие. У вторичноротых – в анальное отверстие, а рот образуется на противоположном конце (хордовые, ланцетник). Инвагинация характерна для ланцетника, а также других организмов (рис. 6).
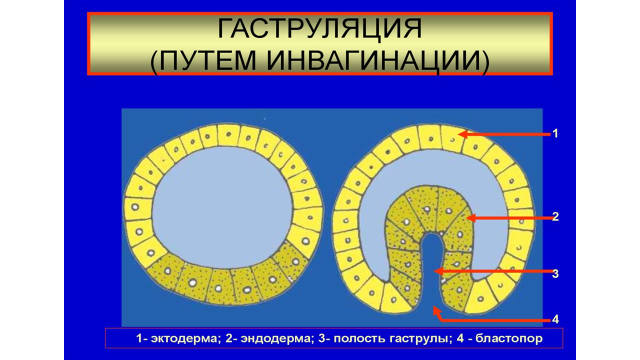
Рис.6. Гаструляция путем инвагинации
Деламинация в чистом виде характерна для кишечнополостных, имеющих бластулу в виде морулы (без полости). При деламинации происходит расслоение клеток бластодермы бластулы на два слоя, расположенных друг над другом. Наружные клетки будут формировать эктодерму. Клетки внутреннего слоя образуют энтодерму (рис. 7).
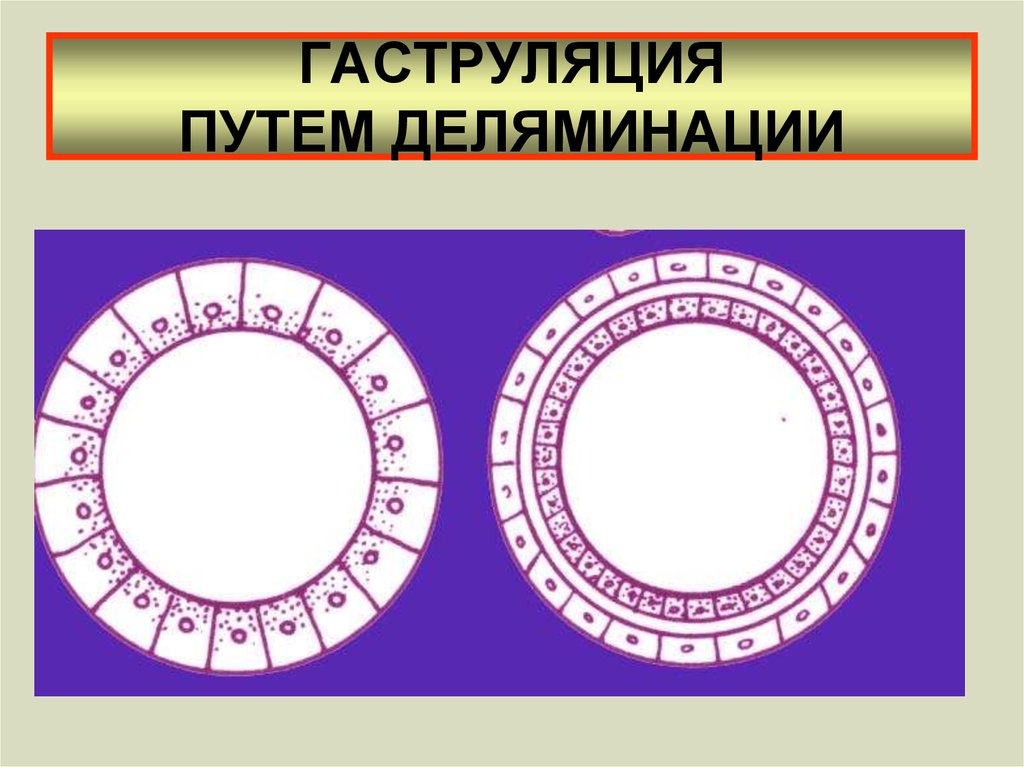
Рис. 7. Гаструляция путем деляминации
При иммиграции отдельные клетки стенки бластулы вычленяются (выселяются) в полость бластоцель. Выселившиеся клетки формируют внутренний слой гаструлы (энтодерму), наружный слой клеток образует эктодерму (рис. 8).
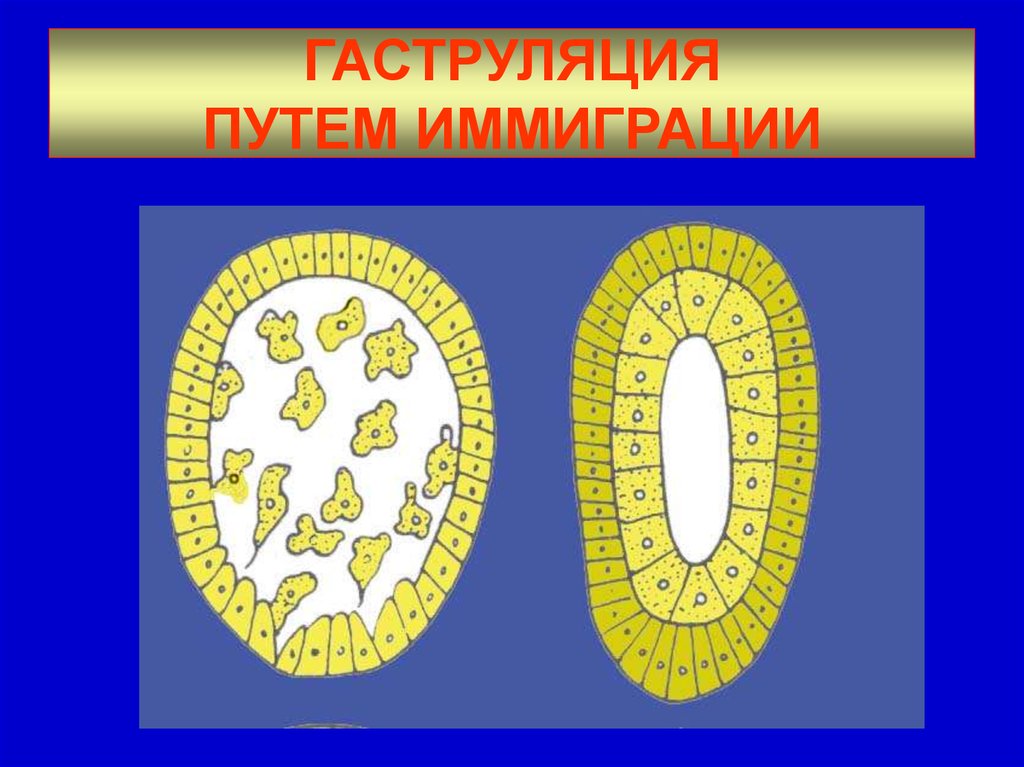
Рис. 8. Гаструляция путем иммиграции
Эпиболия – это перемещение с анимального полюса мелких быстро делящихся клеток (микромеров) в направлении вегетативного полюса и обрастание ими более крупных и менее подвижных клеток вегетативного полюса (макромеры). При этом формируется два слоя клеток – наружный (эктодерма) и внутренний (энтодерма). Такой способ выражен у амфибий (рис. 9).
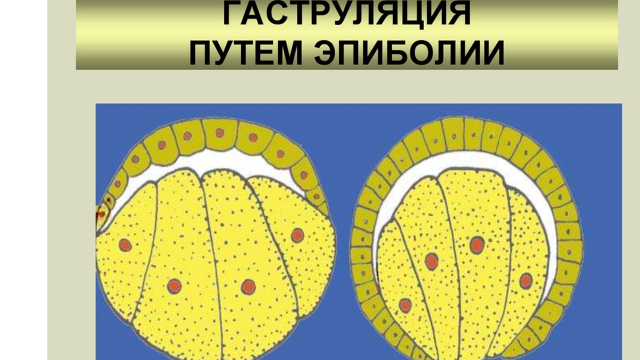
Рис. 9. Гаструляция путем эпиболии
Рассматривая способы гаструляции, надо иметь ввиду, что в чистом виде они редко встречаются, имеет место сочетание этих способов у разных видов. Так у амфибий имеет место инвагинация и эпиболия, у рептилий и птиц - деламинация и иммиграция. У млекопитающих, в том числе человека, гаструляция идет путем деламинации и иммиграции.
Способы закладка мезодермы. Различают два принципиально отличных способа закладки мезодермы (рис. 10). Первый – телобластический, характерен для первичноротых животных. При телобластическом способе мезодерма развивается из двух бластомеров симметрично расположенных в области губ бластопора. При делении этих клеток возникает пара мезодермальных полосок. Позже они подразделяются на сомиты. Принципиально другой способ закладки мезодермы – энтероцельный, свойственен вторичноротым животным. При этом способе мезодерма формируется из клеток энтодермы (участков первичной кишки), которые симметрично впячиваются в полость бластоцель. В последующем эти участки отделяются, дифференцируются в мезодерму и подразделяются на сомиты.
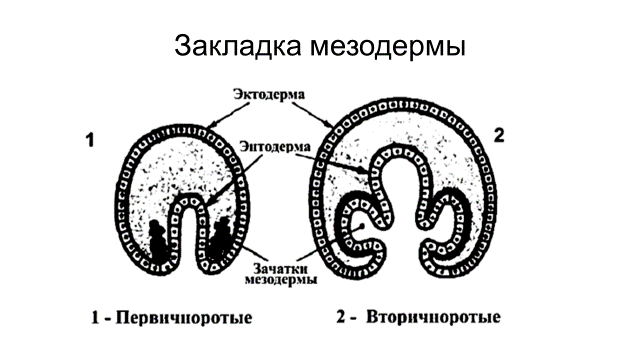
Рис. 10. Телобластический (первичноротые) и энтероцелный (вторичноротые) способы закладки мезодермы
Из зародышевых листков образуются различные ткани, органы и системы органов. Процесс формирования тканей и органов получил название гисто- и органогенез. В процессе формирования органов можно условно выделить две фазы. Первая фаза (начало органогенеза) – нейруляция. Сущность ее состоит в образовании комплекса осевых органов – нервной трубки и хорды. Вторая фаза – это построение всех остальных структур и органов. Остановимся только на нейруляции. Зародыш на стадии нейруляции называется нейрулой (рис. 11).
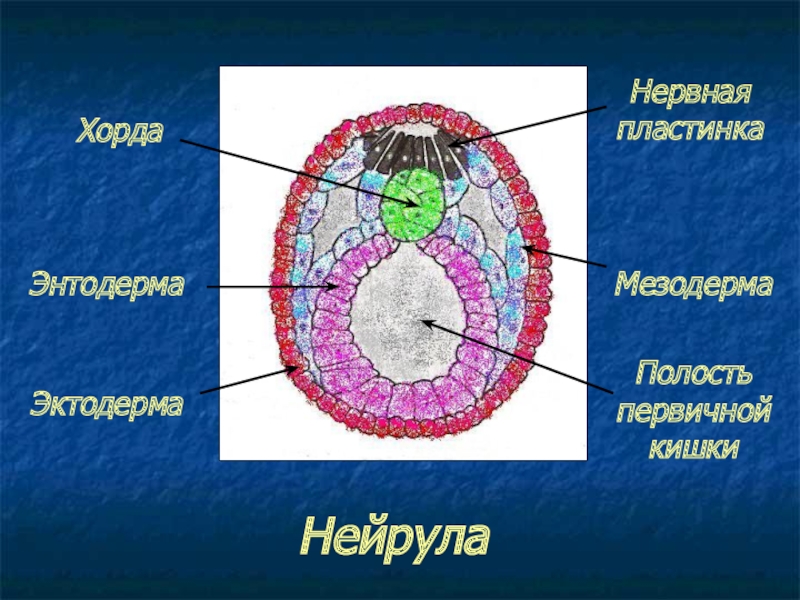
Рис. 11. Нейрула
Образование нервной трубки происходит следующим образом: дорзальная эктодерма гаструлы около срединной линии начинает утолщаться и превращается в нервную пластинку. Вскоре ее боковые края приподнимаются, образуются нервные валики. Нервные валики постепенно сближаются, а нервная пластинка начинает прогибаться вдоль средней линии, образуя углубление нервной пластинки, которое называется нервным желобком. Чуть позже края нервной пластинки смыкаются и формируется нервная трубка, полость которой называется невроцелем. Из переднего отдела нервной трубки в дальнейшем образуется головной мозг, из остальной части – спинной мозг. Параллельно формируется и хорда. Она образуется путем обособления участков первичной кишки по срединной линии. Образуется хорда из зачатка общего с энтодермой и мезодермой. По бокам от образовавшейся хорды лежат обширные участки мезодермы. Вначале мезодерма на каждой стороне представлена единой массой, но вскоре она делится на «блоки» - сомиты. Из них образуются повторяющиеся сегментарные структуры тела позвоночного животного, сомит - сегмент. Сомиты обособляются последовательно один за другим по направлению от головы к хвосту.
Не рассматривая более позднюю фазу органогенеза, тем не менее надо знать какие производные образуются из разных зародышевых листков. Из эктодермы развивается: нервная система, органы чувств, кожный эпидермис, кожные и молочные железы, придатки кожи, эпителий передней и задней кишки (эпителий ротовой полости и анального отдела прямой кишки), эмаль зубов. Из энтодермы: эпителий средней кишки (желудка и кишечника), пищеварительные железы (клетки печени и поджелудочной железы), дыхательная система. Из мезодермы: опорно-двигательный аппарат, кровеносная и лимфатическая система, мочеполовая система, соединительная ткань.