

Лекция 6
Хромосомные основы наследственности. Сцепленное наследование. Работы Т. Моргана по сцепленному наследованию. Генетические и цитологические карты хромосом. Механизмы генетического определения пола. Сцепленное с полом наследование.
Хромосомные основы наследственности. Сцепленное наследование
Как отмечалось в главе 3, Г. Мендель впервые высказал идею о существовании наследственных факторов, которые В. Иогансен назвал генами. Однако до начала второго десятилетия прошлого века никто не знал где находятся гены, несмотря на то, что наличие хромосом в ядре клетки было выявлено задолго до того, как В. Вальдейр в 1888 г. присвоил им название хромосомы.
Мендель установил закон о независимом наследовании признаков, но многими исследователями было показано, что действие третьего менделевского закона ограничено, имеет место зависимая (совместная) передача потомству различных признаков. Особое значение в понимании существа этого явления имели работы Т. Моргана и его учеников по изучению сцепленного наследования у мушки D. melanogaster. Опыты Т. Моргана с мушкой дрозофилой пролили свет на явление зависимой передачи потомству различных генов и послужили одним из доказательств хромосомной локализации генов.
В первых своих опытах Т. Морган проводил скрещивание мух серых с длинными крыльями, то есть нормальных по обоим признакам (за эти признаки отвечают доминантные гены: B - серый цвет, V - длинные крылья), с мутантными черными мушками с короткими крыльями (b - черный цвет, v - короткие крылья). Таким образом, было проведено дигибридное скрещивание мух, отличающихся по двум парам признаков (цвету тела и размеру крыльев). В первом поколении были получены исключительно нормальные мухи вследствие доминирования сервой окраски (B) над черной (b) и длинных крыльев (V) над короткими (v). До сих пор полученные результаты соответствовали тому, что следует ожидать на основе закона Г. Менделя о единообразии гибридов первого поколения:
P |
♀BBVV |
x |
♂bbvv |
|
сер. цв., дл. кр. |
|
чер. цв., кор. кр. |
G |
BV |
|
bv |
|
|
|
|
F1 |
|
BbVv |
|
|
|
сер.цв.,кор.кр. |
|
Далее Т. Морган провел анализирующее скрещивание. Он скрестил гибридного самца из первого поколения с рецессивной гомозиготной самкой. На основании закона чистоты гамет и закона независимого распределения хромосом при мейозе дигибридный самец должен дать четыре сорта гамет, а именно: BV, Bv, bV, bv. При анализирующем скрещивании должно, согласно закона Г. Менделя, получиться четыре фенотипических вида потомков в отношении 1:1:1:1. Наряду с исходными родительскими формами (серый цвет - длинные крылья и черный цвет - короткие крылья) должны появиться также мухи черные длиннокрылые и серые с короткими крыльями. Однако Т. Морган не получил данные в подтверждение этого расчета, основанного на законе Г. Менделя. Вместо четырех типов особей при анализирующем скрещивании он получил только два: мух черных с длинными крыльями и нормальных по обоим признакам. Таким образом, при анализирующем скрещивании получились лишь исходные формы, которые были взяты первоначально для скрещивания. Полученный результат позволил Т. Моргану сделать вывод, что гены, ответственные за черный цвет тела и короткие крылья, наследуются вместе. Они сцеплены между собой, так же как и доминантные аллели этих генов, отвечающие за серый цвет и длинные крылья.
Такой результат должен получиться если предположить наличие связи генов с хромосомами и, что гены B и V находятся в одной гомологичной хромосоме, а аллели b и v - в другой гомологичной хромосоме. При этом в силу расхождения гомологов при мейозе гибридный самец может дать только два сорта гамет BV и bv:
F1 |
♂ |
|
B |
|
b |
|
|
|
|
V |
|
v |
|
|
|
|
↓ |
|
↓ |
|
G |
|
BV |
← |
|
→ |
bv |
При сочетании этих гамет в анализирующем скрещивании с одним сортом гамет, дающей рецессивной гомозиготной самкой (bv), может образоваться только два вида потомков. Гаметы bV и Bv не могут возникнуть без нарушения целостности хромосом, они отсутствуют среди половых клеток гибрида. Отсюда совершенно понятно, каким образом в результате анализирующего скрещивания самца-гибрида с рецессивной самкой по обоим генам получаются в потомстве лишь две фенотипические формы вместо ожидаемых четырех.
Схема анализирующего скрещивания при полном сцеплении генов:
P ♂ BbVv x ♀ bbvv
сер. цв., чер. цв.,
дл. кр. кор. кр.
G BV bv bv
F BbVv bbvv
сер. цв., чер. цв.,
дл. кр. кор. кр.
Совместное наследование генов, ограничивающее свободное их комбинирование, называют сцепленным наследованием. Изучение сцепленного наследования и сопоставление с поведением хромосом при гаметогенезе и оплодотворении позволило Т. Моргану доказать хромосомную локализацию генов и тем самым заложить фундамент в создание хромосомной теории наследственности.
После открытия сцепления генов Т. Морганом было установлено, что у дрозофилы имеется 4 группы сцепленных генов. Это было сопоставлено с наличием 4 хромосом в гаплоидном наборе дрозофиллы и сделан вывод, что количество групп сцепления генов равно числу хромосом в гаплоидном наборе. Таким образом, гены локализованные в одной хромосоме и наследуемые вместе составляют группу сцепления генов. Наличие групп сцепления и соответствие их количества числу хромосом в гаплоидном наборе явилось еще одним доказательством роли хромосом в наследственности. Более того, исследование сцепленного наследования генов позволило установить, что определенная группа сцепленных генов связана с определенной хромосомой.
Итак, явление сцепленного наследования генов существует. Однако оказалось, что сцепление генов в хромосоме не абсолютно. Существует неполное сцепление генов, локализованных в одной хромосоме. Неполное сцепление было показано в других опытах Т. Моргана. В этих опытах Т. Морган для анализирующего скрещивания брал не гибридного самца, а гибридную самку.
У гибридных самок полное сцепление генов в большей или меньшей степени нарушается. При анализирующем скрещивании было получено не два, а четыре фенотипических типа потомков. Кроме черных мух с короткими крыльями и мух, имеющих серый цвет и длинные крылья, были получены мухи серые с короткими крыльями и черные с длинными крыльями. Таким образом, у гибридных самок происходит нарушение сцепления генов, ведущее к появлению новых комбинаций генов и появлению новых типов потомков. Нарушение сцепления генов у гибридной самки связано с кроссинговером, при котором происходит обмен генами (например, В и b) между гомологичными хромосомами в профазу мейоза I. Однако кроссинговер происходит только в части клеток при образовании гамет у гибридных самок. В этой связи гибридные самки дают не 2, а 4 сорта гамет:
|
|
|
|
|
||||
F1 |
♀ |
|
B |
|
↔ |
|
b |
|
|
|
|
V |
|
|
|
v |
|
|
|
↓ |
|
↓ |
||||
G |
|
BV |
|
bV |
|
Bv |
|
bv |
|
|
|
|
|
||||
В результате случайного и равновероятного сочетания четырех сортов гамет гибридной самки с одним сортом гамет рецессивного самца в анализирующем скрещивании образуется 4 типа потомков (см. схему скрещивания).
Схема анализирующего скрещивания при неполном сцеплении генов:
P |
|
♀BbVv |
x |
♂bbvv |
||||
|
|
сер. цв., дл. кр. |
|
чер. цв., кор. кр. |
||||
G |
BV Bv bV bv |
|
|
bv |
|
|||
|
|
|
|
|
||||
F |
BbVv |
Bbvv |
bbVv |
bbvv |
||||
|
сер. цв., дл. кр. |
сер. цв., кор. кр. |
чер. цв., дл. кр. |
чер. цв., кор. кр. |
||||
|
41,5% |
8,5% |
8,5% |
41,5% |
||||
Однако соотношение потомства при анализирующем скрещивании получилось не по 25%, как должно было быть для дигибридного анализирующего скрещивания по закону Г. Менделя о независимом наследовании пар аллелей. Черных мух с короткими крыльями и нормальных по обоим признакам было по 41,5%, мух с новыми комбинациями, черных с длинными крыльями и серых с короткими крыльями - по 8,5%. Всего мух с новыми комбинациями признаков получилось 17%. Поскольку эти мухи (17%) появились в силу кроссинговера, их называют кроссоверными особями. Так происходит только у самок, у самца F1 кроссинговера не происходит (биологическая особенность), имеется полное сцепление указанных генов, поэтому гибридный самец дает только два сорта гамет.
Результаты всех этих опытов показывают, что действительно существует сцепление генов и лишь в определенном проценте случаев оно нарушается вследствие кроссинговера (перекреста). При кроссинговере между гомологичными хромосомами осуществляется взаимный обмен идентичными участками. Величина (частота) кроссинговера определяется отношением кроссоверных особей к общему числу потомков анализирующего скрещивания и выражается в %. В данном случае она равна 17%. Величина кроссинговера отражает силу сцепления генов в хромосоме. Чем больше величина кроссинговера, тем меньше сила сцепления между конкретными генами, тем дальше гены расположены друг от друга. Сила сцепления между генами обратно пропорциональна расстоянию между ними (правило Моргана).
Таким образом, Т. Морган установил, что частота кроссинговера показывает относительное расстояние между генами. За единицу расстояния между генами в хромосоме принимают один процент (1%) кроссинговера между ними. Эта единица называется морганидой – М (в зарубежной генетической литературе более часто используют термин сантиморганида - сМ). Если два гена расположены в хромосоме на расстоянии в одну морганиду - это значит, что кроссинговер между ними происходит в 1% делений клеток. Если гены одной группы сцепления расположены на расстоянии 50М и более друг от друга - они наследуются независимо.
На основании многочисленных генетических исследований полного и неполного сцепления генов Т. Морган выдвинул принцип о линейном расположении генов в хромосомах. Он полагал, что гены в хромосоме расположены в линейном порядке друг за другом и занимают там определенное место. На принципе линейного расположения генов в хромосоме и кроссинговере основано построение генетических карт хромосом.
Генетическая карта хромосомы - это схема относительного положения генов в хромосоме, с учетом расстояния между ними. Генетическую карту хромосомы можно представить, как отрезок прямой, на котором обозначен порядок расположения генов и указано расстояние между ними в морганидах (рис. 1).
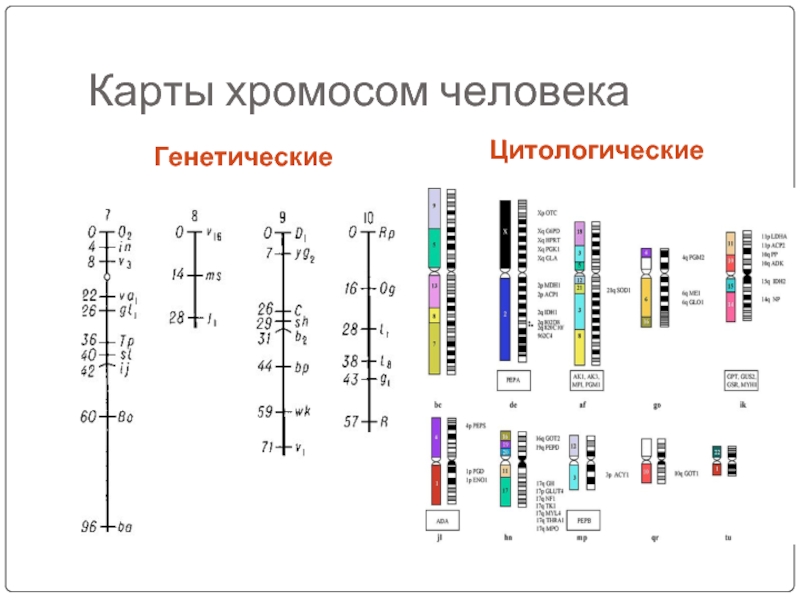
Рис. 21. Генетические и цитологические карты хромосом
Генетическую карту хромосомы по всей ее длине строят постепенно, складывая данные о частоте кроссинговера между двумя последовательно расположенными наиболее близкими генами. Например, если известно, что процент кроссинговера между генами А и С группы сцепления составил 5% и необходимо установить место третьего гена B в той же хромосоме, то необходимо выяснить процент кроссинговера между генами B и генами A и C. Если процент кроссинговера между A и B составляет 6%, то ген B находится на одном из концов хромосом. На основании таких экспериментов в настоящее время генетические карты составлены для многих организмов. Наиболее подробные карты известны для дрозофилы, душистого горошка, кукурузы, мыши, кролика и т.д.
Ведется активно построение генетических карт хромосом человека на основе анализа родословных. В 1911 году впервые была определена хромосомная локализация гена ответственного за цветовую слепоту (дальтонизм) в Х-хромосоме. Хромосомная локализация первого аутосомного гена была определена в 1968 году. Было установлено, что ген, контролирующий антигены крови на поверхности эритроцитов, локализован в хромосоме 1. С тех пор на основании анализа родословных определены группы сцепления для десятков генов человека.
Трудности построения генетических карт хромосом человека по анализу родословных обусловлены тем, что анализируемое потомство малочисленно, поколения сменяются медленно, а подбор пар не подвергается планированию.
Задача картирования хромосом человека оказалась существенно облегченной благодаря внедрению методов работы с соматическими клетками.
В настоящее время картирование хромосом человека проводится путем анализа родословных и генетического исследования гибридных соматических клеток. Для картирования хромосом применяют гибридизацию соматических клеток человека с клетками различных животных, преимущественно с клетками мыши. При культивировании таких гибридных клеток на питательной среде происходит постепенная потеря хромосом человека. Когда в гибридной клетке остается только одна хромосома от кариотипа человека, ее можно идентифицировать путем дифференциального окрашивания. Определив затем содержимое определенного фермента в данной клетке, можно утверждать, что ген для этого фермента расположен в данной хромосоме. В настоящее время определено положение многих сотен генов в хромосомах человека. Подавляющее большинство этих генов локализовано методами генетики соматических клеток.
Составление точных генетических карт хромосом является необходимым условием дальнейшего прогресса теоретической генетики человека. Это имеет исключительное значение для выявления и дифференциальной диагностики наследственных болезней человека, определения носителей генетических сдвигов, которые находятся в бессимптомной фазе, для определения медико-генетического прогноза. В настоящее время не существует принципиальных технических препятствий для получения полной карты генома человека. Однако вследствие огромного размера изучаемой ДНК этот процесс, конечно, займет много времени.
Наряду с генетическим картированием проводится и цитогенетическое картирование хромосом с построением цитологических карт хромосом, представляющих собой схематическое изображение хромосомы с указанием места фактического расположения отдельных генов (рис. 21). Этот метод основан на использовании хромосомных перестроек. Он позволяет прямо сопоставлять результаты генетического анализа расстояний между изучаемыми генами, их взаимного расположения с хромосомными перестройками и на основании этого устанавливать место того или иного гена в хромосоме.
В настоящее время осуществляется составление и физических карт хромосом, характеризующихся в сравнении с генетическими картами большей разрешающей способностью. Расстояние между анализируемыми генами при физическом картировании выражается в парах нуклеотидов. Генетическое расстояние в 1 М соответствует физическому расстоянию между генами примерно в 1 млн. пар нуклеотидов.
Все выше представленные результаты исследований Т. Моргана и его коллег были обобщены в виде хромосомной теории наследственности. Согласно этой теории:
1) гены расположены в хромосомах;
2) гены располагаются в хромосомах в линейном порядке, в определенной последовательности;
3) каждый ген в хромосоме занимает определенное место;
4) каждая хромосома представляет отдельную группу сцепления генов;
5) число групп сцепления генов равно числу хромосом в гаплоидном наборе.
Генетика пола. Сцепленное с полом наследование
Соотношение полов в природе, как правило, 1:1, на 100 рождающихся самцов приходится 100 самок. Такое расщепление соответствует моногибридному расщеплению при анализирующем скрещивании, если один из полов аналогичен гетерозиготному организму (Аа), а другой гомозиготному (аа).
Если пол наследуется по такому принципу, как и другие признаки, то следует предположить, что один пол, например женский, должен быть «гомозиготным», а мужской «гетерозиготным», или наоборот. Тогда действительно в потомстве расщепление по полу оказывается равным 1:1. Такая догадка была высказана еще Г. Менделем. В последствии в исследованиях К. Коренса (1907), Л. Донкастера (1906) эта догадка получила убедительное подтверждение. Однако одни генетические опыты не могли дать решающего доказательства гомо- и гетерогаметности полов, пока полученные в них результаты не были сопоставлены с цитологическими фактами. Было показано, что определение пола зависит от сочетания специальных, получивших название половых, хромосом. Особый вклад в этот раздел цитогенетики внес американский ученый Э. Вильсон. Исследования по цитологии половых различий многих насекомых, многих двудольных растений, большинства позвоночных и человека показали, что один пол имеет две одинаковые Х-хромосомы, другой пол имеет гетероморфную пару из Х- и У-хромосом (ХУ). Пол, который имеет две одинаковые половые хромосомы (ХХ) получил название гомогаметного, так как он образует гаметы только одного сорта с Х-хромосомой. Пол, который имеет гетероморфную пару (ХУ) называют гетерогаметным, так как он дает в равном количестве два вида гамет (50% с Х и 50% с У). У этих видов организмов набор в виде пары ХУ характеризует самцов, а набор хромосом ХХ характеризует самок. Впервые такой состав половых хромосом был обнаружен у клопа Lygaeus turcicus. Поэтому этот способ определения пола носит название тип Lygaeus или тип ХУ.
Рассмотрим, как определяется пол у организмов, относящихся к типу Lygaeus. Принимая во внимание, что аутосомы в норме не влияют на определение пола, схему определения пола при типе ХУ можно представить:
P |
♀XX |
x |
♂XY |
||||
|
|
|
|
||||
G |
|
X |
|
|
X |
|
Y |
|
|
|
|
||||
F1 |
♀XX |
|
♂XY |
||||
|
1 |
: |
1 |
||||
Из схемы видно, что соотношение полов будет 1:1.
У ряда видов организмов гетероморфность в паре половых хромосом выражается не в присутствии У-хромосомы, а в ее отсутствии. В этом случае один пол имеет набор ХО - самцы, а другой ХХ - самки. Впервые такой состав половых хромосом был обнаружен у клопа Protenor, поэтому этот способ определения пола носит название тип Protenor или тип ХО. Такой тип обнаружен у многих насекомых (жуки, пауки, чешуекрылые). Схематически систему определения пола при типе Protenor можно представить следующим образом:
P |
♀XX |
x |
♂XO |
||||
|
|
|
|
||||
G |
X |
|
X |
|
X |
|
O |
|
|
|
|
||||
F1 |
♀XX |
|
♂XO |
||||
|
1 |
: |
1 |
||||
Достаточно распространен третий способ определения пола, при котором гетерогаметность присуща женским особям, а гомогаметность - мужским. Первоначально этот метод был открыт у бабочки Крыжовенной пяденицы. Встречается он у многих видов птиц, пресмыкающихся, некоторых рыб, земноводных. При гетерогаметности женского пола описаны подтипы определения пола, аналогичные Lygaeus и Protenor: а) самки ХУ, самцы ХХ и б) самки ХО и самцы ХХ.
Схемы определения пола в этих случаях принципиально не отличаются от типа ХУ и типа ХО:
P |
♀XY |
x |
♂XX |
|
|
P |
♀XO |
x |
♂XX |
||||||||||||||||
|
|
|
|
|
|
|
|
|
|
||||||||||||||||
G |
X |
|
Y |
|
|
X |
|
|
|
G |
X |
|
O |
|
|
X |
|
||||||||
|
|
|
|
|
|
|
|
|
|
||||||||||||||||
F1 |
♀XY |
|
♂XX |
|
|
F1 |
♀XO |
|
♂XX |
||||||||||||||||
|
1 |
: |
1 |
|
|
|
1 |
: |
1 |
||||||||||||||||
Совершенно другой механизм определения пола, называемый гаплодиплоидией, широко распространен у пчел и муравьев. У этих организмов нет половых хромосом: самки - это диплоидные особи, а самцы (трутни) - гаплоидные. Самки развиваются из оплодотворенных яиц, а из неоплодотворенных развиваются трутни. У трутней, таким образом, нет отцов, хотя у них есть деды по материнской линии. В процессе сперматогенеза у трутней не происходит редукции хромосом. Из оплодотворенной яйцеклетки может развиваться либо «матка» - крупная, способная к размножению самка, либо стерильная рабочая самка. Это зависит от условий вскармливания личинки рабочими пчелами. Следует отметить, что гаплоидность у трутней сохраняется почти всегда только в клетках зародышевого пула, тогда как в соматических клетках восстанавливается диплоидность. Такое вторичное восстановление диплоидности приводит к полной гомозиготности клеток тканей. Поэтому рецессивные гены проявляют свой отрицательный эффект еще на ранних стадиях развития трутня, понижая его жизнеспособность или вызывая летальное действие. За счет гаплоидии популяция пчел как бы очищается от действия вредных генов.
Таким образом, определение пола будущего организма происходит в момент оплодотворения при помощи хромосомного механизма. Главным фактором во многих случаях, который сдвигает фенотип в мужскую сторону, является У-хромосома. Однако формирование конкретного фенотипа происходит не только с учетом особенностей генотипа, но и в зависимости от условий среды. Насколько сильно оказывают влияние условия развития организма на формирование конкретного фенотипа свидетельствует тот факт, что возможно отклонение фенотипа организма в сторону противоположного пола, несмотря на соответствующее сочетание половых хромосом, то есть возможно переопределение пола. Показано переопределение пола в жизненном цикле некоторых рыб, моллюсков в зависимости от стадии жизненного цикла и условий среды. При этом происходит развитие не только соответствующих вторичных половых признаков, но и первичных (соответствующих половых желез). Возможность переопределения пола указывает на чрезвычайно важную роль условий развития организма, факторов внешней среды на процесс формирования фенотипа. Например, у морского червя Bonellia особи, которые в личиночной стадии остаются свободно плавающими, становятся самками, а личинки, прикрепившиеся к телу взрослой самки, превращаются в самцов в результате маскулинизирующего действия гормона, выделяемого самкой.
С полом наследуются многие признаки. Наследование признаков, гены которых находятся в Х- и У-хромосомах, называют наследованием сцепленным с половыми хромосомами или сцепленным с полом наследованием. Это явление было открыто Т. Морганом у дрозофилы и явилось еще одним доказательством роли хромосом в наследственности. У человека целый ряд признаков также наследуется сцепленно с полом, в том числе и патологических.
Признаки, которые наследуются через У-хромосому могут быть только у лиц мужского пола, а наследуемые через Х-хромосому как у женского, так и мужского. Генетическими исследованиями установлено, что У-хромосома человека существенно отличается от Х-хромосомы. Она содержит меньше генов по сравнению с Х-хромосомой, а гены, которые обеспечивают жизнеспособность организма, в У-хромосоме фактически отсутствуют. В Х- хромосоме содержится более 1000 генов, У-хромосома содержит 89 генов, кодирующих белки. Х- и У-хромосомы имеют небольшое количество гомологичных локусов (рис. 2). Поэтому почти все локализованные в Х-хромосоме гены, даже рецессивные, проявляются в фенотипе мужского организма. Такие организмы получили название гемизиготные.
|
|
негомологичный участок Y-хромосомы |
Y→ |
|
|
|
|
|
|
|
←Х |
негомологичный участок Х-хромосомы |
гомологичные участки |
|
Рис. 2. Гомологичные и негомологичные участки Х- и У-хромосом
У человека выделяют три типа наследования, сцепленного с полом: тип Х, тип У, тип ХУ.
Тип Х. При этом типе наследуемые гены локализованы в негомологичных участках Х-хромосомы и не имеют аллелей в У-хромосоме. По такому типу наследуются такие патологические состояния у человека как гемофилия, дальтонизм, близорукость, мышечная атрофия Говера и др. Наследуют эти признаки женщины и мужчины. Однако если признак рецессивный, то мужчины наследуют признак чаще, а если признак доминантный, то чаще наследуют этот признак женщины. Например, гемофилией болеют почти всегда мужчины. Заболевание женщин маловероятно, так как для этого они должны иметь соответствующий рецессивный ген в обеих Х-хромосомах. Одну из этих хромосом женщины должны получить от отца больного гемофилией. Однако больные мужчины редко доживают до репродуктивного возраста. В то же время сыновья женщины-носительницы гемофильного гена непременно получат этот ген и заболеют.
P |
♀ XHXh |
x |
♂XHY |
||||||
|
|
|
|
||||||
G |
XH |
|
Xh |
|
XH |
|
Y |
||
|
|
|
|
||||||
F1 |
XHXH |
XHY |
XHXh |
XhY |
|||||
|
Норма |
Норма |
Носитель |
Гемофилия |
|||||
Аналогично наследуется дальтонизм, неспособность различать красный и зеленый цвет.
Тип У. Наследуемые гены локализованы в негомологичных участках У-хромосомы и не имеют аллелей в Х-хромосоме. По такому типу наследования гены, локализованные в У-хромосоме, передаются прямо от отца к сыну, причем в каждом поколении. Так наследуется, в частности признак «волосатых ушей» (гипертрихоз края ушной раковины). Признак проявляется в том, что на ушной раковине вырастает пучок длинных жестких волос. Такой тип наследования получил название голандрическое наследование, а гены - голандрические гены.
Тип ХУ. При этом типе наследуемые гены локализованы в гомологичных участках Х- и У- хромосомы. Подобные гены считаются частично сцепленными с полом. У человека известно несколько патологических состояний, наследуемых по такому типу. К ним относится: полная цветная слепота, пигментная ксеродермия, пигментный ретинит, спастический паралич, волчья пасть, заячья губа и др.