

Лекция 12
Хромосомные мутации. Механизмы их обусловливающие. Фенотипический эффект.
Хромосомные мутации (хромосомные перестройки, хромосомные
аберрации) представляют собой структурные перестройки, в результате которых утрачиваются, добавляются или перестраиваются участки одной или нескольких хромосом. При этом участки хромосом, содержащие блоки генов, могут перемещаться внутри одной хромосомы или между разными хромосомами. В этой связи выделяют внутрихромосомные и межхромосомные перестройки. При внутрихромосомных перестройках изменения происходят в пределах одной хромосомы. Сюда относятся: делеция, дупликация, инверсия, кольцевые хромосомы, изохромосомы. К межхромосомным перестройкам относят обмен участками между разными хромосомами, что вызывает изменение распределения генов по группам сцепления (транслокация).
Хромосомные мутации широко представлены у растений, животных, известны они и у человека. Происходят они как в половых, так и соматических клетках. Если хромосомные перестройки происходят в половых клетках, то гаметы будут нести хромосомные аберрации, которые наследуются в потомстве. Аберрации могут возникать как спонтанно, так и индуцировано. Наиболее часто в роли мутагенов выступают ионизирующая радиация, химические вещества.
В настоящее время выдвигается два объяснения механизма возникновения хромосомных мутаций. Во-первых, часто такие хромосомные перестройки связаны с одним или несколькими разрывами хромосом с последующей потерей или перегруппировкой генетического материала. Разрывы могут происходить в одной или одновременно в двух хромосомах. Разрывы могут возникать во время интерфазы как до, так и после репликации ДНК. Если разрыв происходит до репликации, повреждение будет обнаруживаться в обеих хроматидах (изохроматидный разрыв). Если разрыв произойдет после репликации, поврежденной окажется только одна хроматида (хроматидный разрыв). Концы хромосомных фрагментов при разрывах могут воссоединиться в точках разрыва других хромосом как гомологичных, так и негомологичных, но при условии, что разрывы происходят в хромосомах одновременно и достаточно близко друг к другу. Это приводит к образованию хромосомных перестроек различного вида. Хромосомные разрывы могут быть как спонтанными, так и индуцированными химическими веществами, радиацией. Во-вторых, хромосомные перестройки могут возникать и в результате нарушения процессов кроссинговера, в частности при неравном кроссинговере.
Говоря о хромосомных мутациях в целом, не разделяя их, следует отметить, что они могут быть сбалансированными и несбалансированными. При сбалансированных мутациях баланс генов не нарушен, система генотипа не изменена и фенотипических проявлений, как правило, нет. При несбалансированных – баланс генов нарушен, изменена система генотипа, что приводит к фенотипическим изменениям. У человека при разных видах несбалансированных аберраций имеет место совпадение многих признаков и симптомов. Их характеризует комплекс физических и психических недостатков: резкая задержка развития, умственная отсталость, дефекты развития со стороны наружных и внутренних органов. В тоже время надо иметь ввиду, что хромосомные аберрации играли важную роль в эволюции генома и хромосом. На их основе осуществлялась эволюция видовых кариотипов.
Делеция (или нехватка). Это аберрация, которая связана с потерей участка хромосомы. Если происходит один разрыв и теряется концевой участок – это терминальная (концевая) делеция. Если два разрыва в хромосоме и выпадает срединный участок, а концы соединяются - это интерстициальная (интеркалярная) делеция. Если хромосома с делецией имеет центромеру, то она передается при делении клеток аналогично другим хромосомам. Фрагмент хромосомы без центромеры обычно утрачивается в ходе клеточных делений.
Делеции могут возникать при неравном кроссинговере. Когда в соседних участках хромосомы оказываются похожие последовательности ДНК, то конъюгация гомологов может произойти неправильно. Неравный кроссинговер в таких неправильно конъюгировавших участках хромосом приводит к образованию гамет с дупликацией или делецией (нехваткой) (рис. 1).
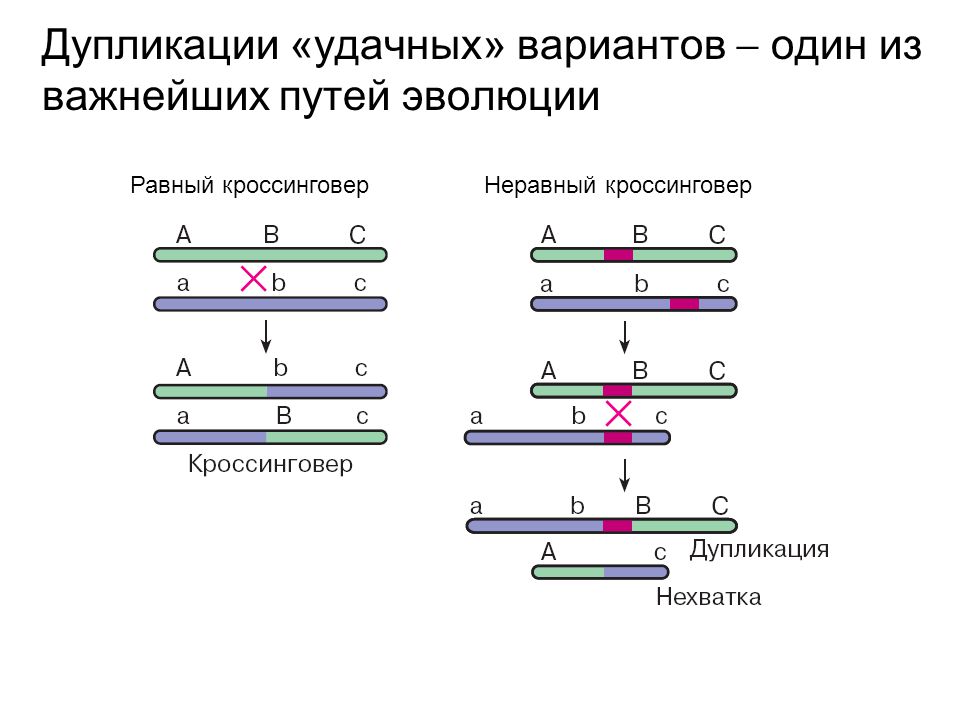
Рис. 1. Схема неравного кроссинговера
Делеции могут быть большими и малыми, могут быть микроделеции, которые очень трудно обнаружить. В гомозиготном состоянии, когда теряются гомологичные участки в обеих гомологичных хромосомах, делеции обычно летальны (а также в гемизиготном состоянии, если делеция произошла в Х-хромосоме). Небольшие делеции сохраняются и передаются по наследству через гетерозигот. При этом у гетерозигот имеет место нехватка по одной хромосоме гомологичной пары. Цитологически делеции можно выявить по появлению петли при конъюгации гомологичных хромосом в профазе мейоза I. Синапсис между нормальной гомологичной хромосомой и другой хромосомой пары, несущей интерстициальную делецию, приводит к появлению петли на первой хромосоме, компенсирующей делетированный участок.
Классическим примером делеции является мутация у дрозофилы, которая в своё время получила название мутация Notch - вырезка на крыле. Эта мутация сцеплена с полом и в гетерозиготном состоянии обуславливает развитие зазубренности на крыле, являясь доминантной. Мутация Notch связана с интерстициальной делецией, локализованной на конце Х-хромосомы. В гомозиготном состоянии у самок и в гемизиготном состоянии у самцов эта делеция приводит к летальности. При изучении этой мутации были обнаружены два генетических явления: первое – явление псевдодоминирования, второе – отсутствие кроссинговера на участке нехватки. Явление псевдодоминировния состоит в том, что у гетерозигот по этой делеции проявляются рецессивные аллели ряда генов, локализованных в гомологичной хромосоме напротив нехватки. Анализ показал, что в случае мутации Notch, выпавший участок хромосомы содержит ряд генов, в том числе и аллель Whitе, обеспечивающий синтез пигмента в глазах. Поэтому у гетерозиготных самок, несущих в одной гомологичной хромосоме нехватку, а в другой рецессивный аллель белоглазия, самки оказались белоглазыми. Такое явление получило название псевдодоминирования, поскольку у особей, имевших рецессивный аллель в гетерозиготном состоянии, тем не менее, проявляются определяемые им признаки. Это связано с тем, что нормальный аллель в другой хромосоме утерян при возникновении делеции. В результате измерения длины хромосомы генетическими методами с помощью кроссинговера показано, что хромосома с делецией короче нормальной. Отсутствие части хромосомы в результате мутации приводит к тому, что в этом месте не происходит кроссинговер.
У человека делеции обнаружены по разным хромосомам. Для проявления делеций в виде тяжелых симптомов она должна захватывать довольно протяженный участок хромосомы. Если делеция приводит к утрате многих генов, она становится летальной, и такие аберрации обычно не доступны для исследования. Наиболее хорошо изучена у человека делеция короткого плеча 4 и 5 хромосомы.
Делеция короткого плеча хромосомы 5 (синдром кошачьего крика). Это заболевание ассоциировано с потерей части короткого плеча хромосомы 5. Частота синдрома среди новорожденных равна 1: 45000. У детей с данной хромосомной аномалией необычный плач, напоминающий мяуканье кошки. У больных отмечается микроцефалия, лунообразное лицо, широкая переносица, деформированные ушные раковины и другие изменения мозгового черепа и лица. Кроме того, имеются пороки со стороны внутренних органов и высокой степени слабоумие. Обычно носители данных делеций погибают в младенческом возрасте или в раннем детстве, но некоторые доживают до взрослого возраста.
Делеция короткого плеча хромосомы 4 (синдром Вольфа-Хиршхорна). Этот синдром ассоциирован с потерей части короткого плеча хромосомы 4. Частота рождения детей с синдромом Вольфа-Хиршхорна 1: 20000 – 1: 50000. Синдром характеризуется многочисленными врожденными пороками с последующей резкой задержкой физического и психомоторного развития. У таких больных характерные черты лица и черепа: высокий лоб, микроцефалия, клювовидный нос, выступающие глаза, крупные оттопыренные уши, расщелины губы и неба и др. Жизнеспособность детей резко снижена. Большинство умирают в возрасте до 1 года.
Дупликация. Дупликацией называется присутствие одного и того же участка хромосомы в одной хромосоме или разных негомологичных хромосомах более чем в одном экземпляре. Дуплицированные участки могут быть расположены друг за другом, то есть тандемно. Последовательности повторов могут быть взаимно противоположными. Такую тандемную дупликацию называют обращенной.
ABCDEF → ABCDCDEF или ABCDDCEF
Многие дупликации, как и делеции, возникают в результате разрывов в хромосомах. Дупликации, как и делеции, могут возникать при неравном кроссинговере. Появляются дупликации и в результате ошибки репликации в интерфазе. На цитологическом препарате гетерозиготность по дупликациям приводит к образованию петель при конъюгации хромосом во время мейоза, аналогично как у гетерозигот по делециям.
Дупликации могут обладать фенотипическими проявлениями. Наиболее известным примером дупликации служит мутация Bar в Х-хромосоме Dr. melanogaster. Мутация Bar возникает вследствие дупликации маленького участка Х-хромосомы. У самок, гетерозиготных по Bar, вместо овальных глаз нормального размера имеются узкие щелевидные глаза. У гомозигот и гемизигот по Bar глаза еще меньше.
Иногда дупликации выявляются благодаря тому, что у особи, гомозиготной по рецессивному аллелю, рецессивный признак тем не менее не проявляется. Этот факт объясняется тем, что соответствующий доминантный аллель содержится в дуплицированном участке хромосомы.
У человека дупликации могут быть по разным хромосомам. Сейчас не осталось ни одной аутосомы человека, по которой не было бы обнаружено того или иного числа частичных дупликаций. Выявляются дупликации у гетерозигот. Дупликации менее вредны, в сравнении с делециями, хотя, если значительно увеличивается количество наследственного материала - это приводит к развитию патологии. В тоже время явление дупликации наследственного материала у организмов важно с эволюционной точки зрения. Будучи в двойном числе, дуплицированные гены структурно и функционально могут преобразоваться. Это служит источником для увеличения объема генетической информации.
Инверсия. Инверсией называют поворот на 180º отдельных участков хромосомы. При этом ни число хромосом, ни число генов в каждой хромосоме не меняется. Происходит лишь изменение порядка расположения генов в хромосоме. Если последовательность генов в исходной хромосоме обозначить ABCDEF и инверсии подвергся участок ВСD, то в новой хромосоме гены будут расположены в последовательности ADCBEF. Инверсии возникают вследствие разрывов в двух точках хромосомы с последующим встраиванием инвертированного фрагмента. Инверсии могут быть одиночными и сложными. В последнем случае последовательно возникают несколько инверсий, что приводит к заметной перестройке блоков генов. Для характеристики инверсий существенно их положение по отношению к центромере. Если разрывы происходят по одну сторону от центромеры и переворачивается на 180º образующийся фрагмент, такую инверсию называют парацентрической. При этом конфигурация хромосомы не изменяется. Если разрывы происходят по обе стороны центромеры и центромера входит в состав инвертированного участка, такую инверсию называют перицентрической. Если концы перевёрнутого участка расположены не симметрично относительно центромеры, это приводит к изменению морфологии (конфигурации) хромосомы.
У гетерозигот по инверсиям при анализе цитологических препаратов хромосом, находящихся на стадии пахитены мейоза, выявляются петли, которые необходимы для конъюгации гомологичных хромосом. Наличие таких петель указывает на гетерозиготность по инверсиям. Инверсии не сохраняются в гомозиготном состоянии, их обнаруживают обычно у гетерозигот. Однако у гетерозигот обычно инверсии не влияют заметным образом на фенотип, они сбалансированы. В тоже время гетерозиготные по инверсиям организмы, как правило, бывают стерильны, поскольку половина образующихся при кроссинговере гамет не способна к образованию жизнеспособных зигот. Гетерозиготы по инверсиям не так уж редки в популяциях людей.
Кольцевые хромосомы. Чаще такие мутации индуцированные. Особенно образуется много кольцевых хромосом при действии ионизирующей радиации. Образование кольцевых хромосом связано с двумя разрывами. При этом обычно утрачиваются обе теломеры (концы хромосом), а открытые концы соединяются друг с другом, формируется центрическое кольцо (рис. 2). Центрическое кольцо передаётся следующим поколениям клеток и организмов.
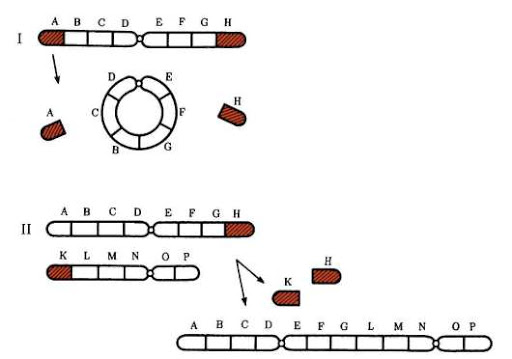
Рис. 2. Формирование центрических кольцевых хромосом
Когда кольцевая хромосома образуется из срединного участка хромосомы, лишенного центромеры, возникает ацентрическая кольцевая хромосома. Как всякое ацентрическое образование хромосомы, ацентрическая кольцевая хромосома при первом же митозе выпадает из ядра в цитоплазму и разрушается. Центрические кольца тоже часто нестабильны как в митозе, так и в мейозе, и часто утрачиваются. Тем не менее кольца описаны почти для всех хромосом человека. В силу утраты наследственного материала при их формировании, они сопровождаются задержкой физического и психического развития и морфологическими аномалиями у гетерозигот.
Изохромосомы. Разрыв хромосомы может происходить через центромеру. Каждое плечо, разъединенное после репликации, имеет две сестринские хроматиды, соединенные оставшейся частью центромеры. Сестринские хроматиды одного и того же плеча становятся плечами одной хромосомы. Со следующего митоза эта хромосома начинает реплицироваться и передаваться из клетки в клетку как самостоятельная единица наряду с остальными хромосомами. Такие хромосомы называют изохромосомами, у них одинаковые по набору генов плечи. Встречаются изохромосомы у человека по короткому или длинному плечу. Наличие изохромосом у человека вызывает хромосомную патологию, поскольку - это одновременно и частичная моносомия (по отсутствующему плечу), и частичная трисомия (по присутствующему плечу). Наиболее часто встречаются изохромосомы Х-хромосомы. Изохромосома длинного плеча Х-хромосомы вызывает синдром Шерешевского-Тернера, поскольку эта хромосома всегда инактивирована, а активна только одна нормальная Х-хромосома.
Транслокация. Перемещение участка одной хромосомы на другую называется транслокацией. К наиболее распространённому типу транслокаций относятся взаимные или реципрокные транслокации. Реципрокные транслокации могут быть между разными хромосомами как гомологичными (но это не кроссинговер), так и негомологичными. При этом происходит обмен участками между гомологичными и негомологичными хромосомами.
Последствия реципрокных транслокаций сходны с последствиями инверсий. Если в результате реципрокных транслокаций происходит рекомбинация генетического материала, но без его утраты в вовлеченных хромосомах, такие транслокации являются сбалансированными и существенно не влияют на жизнеспособность носителей этой аберрации. Однако, как и при инверсии, у гетерозигот с такими транслокациями образуются гаметы, несущие аберрации (дефектные гаметы). При оплодотворении таких аберрантных гамет обычно формируются нежизнеспособные зиготы по аберрациям, а выживает только 50% зигот, гетерозиготных по реципрокной транслокации. Поэтому у носителей транслокаций наблюдается полустерильность. У гетерозигот по реципрокным транслокациям в профазе мейоза наблюдается характерная структура – крест (крестообразный синапсис). Его появление связано с тем, что гомологичные участки, оказавшиеся в разных хромосомах, притягиваются при конъюгации.
Реципрокные транслокации могут быть у человека по разным хромосомам. Довольно частым типом транслокации у человека являются робертсоновские транслокации или центрические слияния. Их формирование связано с утратой в результате разрывов коротких плечей двух акроцентрических хромосом и одной центромеры. Происходит слияние длинного плеча одной хромосомы с длинным плечом другой, при котором формируется одна большая субметацентрическая или метацентрическая хромосома. В 4-5% случаев с такой транслокацией связано развитие синдрома Дауна (транслокационная форма). Синдром связан с центрическим слиянием 14 и 21 хромосом (14/21). Один из родителей в этом случае несёт сбалансированную робертсоновскую транлокацию (14/21) (рис. 3). Его кариотип будет:

Рисунок 3. Сбалансированная робертсоновская транслокация
Такая транслокация не проявляется в фенотипе носителей, но в результате мейоза после конъюгации хромосом у них формируется ¼ гамет с двумя хромосомами 21 - одной нормальной и одной транслоцированной на хромосому 14 (14/21,21). После слияния при оплодотворении таких гамет с нормальной гаметой зигота будет нести три хромосомы 21: 21/14,21 + 14,21 = 21/14,14,21,21, что обусловливает развитие заболевания (рис. 4).
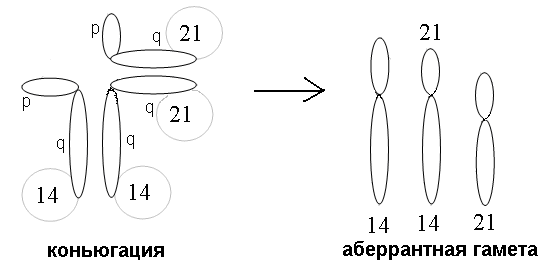
Рис. 4. Образование аберрантных гамет при коньюгации хромосом с сбалансированной робертсоновской транслокацией
Другие типы гамет после оплодотворения нормальными гаметами дают потомство либо с нормальным кариотипом, либо с сбалансированной транслокацией.
Транспозиция. Транспозицией называют перемещение участка хромосомы либо внутри той же хромосомы, либо в другую хромосому. В отличие от реципрокных транслокаций они не имеют взаимного характера. Интересный и важный класс транспозиций связан с функционированием подвижных (мобильных, транспозирующихся) генетических элементов, которые иногда называют транспозонами.
У генетиков в своё время существовало твёрдое правило (установлено Т. Морганом), что каждый ген занимает определённое и постоянное место в хромосоме. В сороковые годы прошлого столетия американская исследовательница Барбара Мак-Клинтон экспериментально показала, что существуют участки хромосом, способные путешествовать по хромосомам, переходить из одного участка генома на другой. Ею был описан первый подвижный генетический элемент кукурузы. В своих исследованиях Барбара Мак-Клинтон использовала гибридологический метод. Однако в то время ее работы не встретили поддержки, поскольку противоречили убеждениям о постоянстве генома. В конце семидесятых годов, когда методами молекулярной биологии было подтверждено открытие Барбары Мак-Клинтон о наличии мобильных генетических элементов, она была представлена к Нобелевской премии, которую получила в 1983 г.
К настоящему времени мобильные генетические элементы (МГЭ) найдены у всех организмов, в которых их искали ( бактерий, дрозофилы, мышей, человека и др.). Обнаружение подвижных генетический элементов как у прокариот, так и эукариот свидетельствует о том, что их присутствие является общим свойством всех организмов. Представлены МГЭ, как правило, многими копиями в геноме. По своему строению МГЭ представляют последовательности нуклеотидов, которые различаются размером, но имеют некоторые общие черты строения. Мобильные элементы обычно имеют на обоих концах одинаковые последовательности (повторы). Это могут быть длинные цепочки нуклеотидов 200-600 оснований или короткие до 30 нуклеотидов. Повторы могут быть прямые и обращенные (расположены в обратном порядке) (рис. 5).
МГЭ
МГЭ
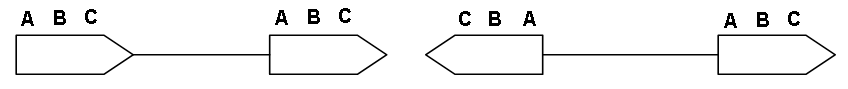
Рис. 5. Прямые и обращенные повторы МГЭ
В 80-х годах прошлого столетия были открыты мобильные диспергированные генетические элементы (МДГЭ) у дрозофилы. Диспергированные - это значит разбросаны по всем хромосомам. Эти элементы состоят обычно из 5-10 тысяч нуклеотидных пар. В геноме дрозофилы открыто около 30 семейств МДГЭ по 10-150 копий в каждом семействе (МДГЭ – 1,2, 3 и т.д.). Все МДГЭ содержат на обоих концах длинные прямые повторы.
Помимо МДГЭ у дрозофилы известны и другие МГЭ, которые устроены по-другому. К этой группе относится семейство Р-элементов генома дрозофилы. Последовательность Р-элемента включает 2900 нуклеотидных пар. На его концах имеются короткие обращенные повторы.
Мобильные генетические элементы имеются и у млекопитающих. Это, так называемые, «вездесущие» короткие последовательности в геноме млекопитающих. Они включают от 100 до 500 нуклеотидных пар. Обнаружены они в геноме мышей и геноме человека. Повторяются эти последовательности в геноме млекопитающих сотни тысяч раз. Представителем таких мобильных элементов у человека является Alu-семейство коротких диспергированных элементов средней длины. Alu-элемент включает 200-300 пар нуклеотидов с числом копий 105-106 на геном. На концах их имеются повторы длиной 7-20 п.н. У млекопитающих в геноме есть семейство и более длинных повторов (не более 6-7 тыс. п.н.). Это средне повторяющиеся последовательности с числом копий у человека 103-105 на геном.
Мобильные генетические элементы представляют собой автономные единицы, в нуклеотидной последовательности которых заключена информация о структуре специализированных белков, обеспечивающих их перемещение. Таким образом, МГЭ сами отвечают за образование аппарата своего перемещения. Предложен ряд возможных механизмов транспозиции.
1. МГЭ могут воспроизводится через обратную транскрипцию. Поток информации может идти не только прямым путем ДНК→РНК, но обратным путем РНК→ДНК с помощью фермента обратной транскриптазы (ревертазы). Сначала информация, кодирующая мобильный ген, с ДНК переписывается на молекулу РНК (прямая транскрипция), а потом эта РНК служит матрицей, на которой синтезируется полинуклеотидная цепь ДНК мобильного гена (обратная транскрипция). Затем ДНК удваивается и образуется копия гена. Эта ДНК образует кольцевую молекулу вне хромосомы, которая встраивается в самые различные участки на разных хромосомах, в то время как исходный МГЭ сохраняет свою прежнюю локализацию. С помощью обратной транскрипции перемещаются МДГЭ дрозофилы.
2. Возможен и другой механизм с участием ферментов – транспозаз. Эти ферменты вырезают мобильные генетические элементы и встраивают их в другое место. Так происходит перемещение Р-элемента дрозофилы.
Исследования показали, что МГЭ перемещаются весьма редко, с частотой не более 10-5 на поколение. Однако в определённых условиях частота перемещения может резко возрастать.
Мобильные генетические элементы - это ДНК, которая не несёт никаких полезных функций в клетке. Они несут информацию только о белках, которые нужны им для перемещения по геному. Каких-то других белков они не кодируют. В тоже время эффект действия МГЭ на жизнь клетки может быть разным.
1. Они могут попасть в кодирующую часть какого-либо гена и изменить генетическую информацию, которую имеет ген, что приведет к генной мутации.
2. МГЭ могут встраиваться рядом с геном и изменять его функцию.
3. Есть определенная связь мобильных элементов и рака. МГЭ могут внедряться рядом или внутрь онкогена и активировать его, что ведет к развитию опухоли. Сейчас уже известны некоторые опухоли, возникновение которых связано с перемещением мобильных элементов.
4. МГЭ могут захватывать с собой соседние участки ДНК и вызывать разные хромосомные перестройки. Они могут вызывать инверсию хромосом, становиться фактором для неравного кроссинговера и т.д.
В то же время с эволюционной точки зрения МГЭ могли быть важным механизмом создания внутривидовой изменчивости хромосомных структур, что связано с их способностью вызывать самые различные хромосомные перестройки.