

Лекция 10
Изменчивость. Формы изменчивости: фенотипическая и генотипическая. Фенотипическая изменчивость, ее характеристика. Комбинативная изменчивость. Мутационная изменчивость. Классификация мутаций. Генные мутации.
Изменчивостью называют способность живых организмов приобретать новые признаки и свойства. Изменчивость определяет разнообразие в строении и функциях организмов, относящихся к одному виду. Важной характеристикой изменчивости является то, что она отражает взаимодействие организма с внешней средой.
По причинам возникновения, по механизму развития изменчивость может быть результатом различных процессов, но с генетической точки зрения изменчивость представляет собой результат реакции генотипа в процессе индивидуального развития организма на условия внутренней и внешней среды. В силу того, что разнообразны причины и механизмы развития изменчивости, выделяют и различные ее формы.
|
Изменчивость |
|
|
||||
|
|
|
|
|
|||
Фенотипическая (модификационная) |
|
Генотипическая |
|
||||
|
|
|
|
|
|||
|
Комбинативная |
|
Мутационная |
||||
Фенотипическая изменчивость
Каждый организм развивается и обитает в определенных условиях, испытывая на себе действие факторов внешней среды: колебания температуры, освещенности, влажности и др. Под влиянием условий среды фенотипические признаки организмов могут меняться. Эти изменения фенотипа и составляют сущность фенотипической изменчивости. На действие определенного фактора внешней среды все организмы биологического вида реагируют специфически и одинаково. Реакция (в форме изменения признака) оказывается сходной у всех особей данного вида. В этой связи фенотипическая изменчивость характеризуется массовым характером изменений, затрагивающих большинство особей в популяции. Поэтому фенотипическую изменчивость называют групповой (определенной по Ч. Дарвину). Фенотипические изменения, которые возникают под влиянием условий среды, называются модификациями. Степень варьирования фенотипических изменений (модификаций) зависит от индивидуальных особенностей генотипа и может колебаться в определенных пределах. Диапазон изменчивости, в пределах которого в зависимости от условий среды один и тот же генотип способен давать различные фенотипы, получил название нормы реакции. Таким образом, норму реакции можно рассматривать как предел модификационной изменчивости. В зависимости от признака норма реакции может быть узкой и широкой. При узкой норме реакции имеет место фактически одинаковое состояние признака в широком спектре условий среды. При широкой норме реакции признак отличается значительной изменчивостью, вариабельностью проявления в зависимости от условий среды.
Норма реакции специфична для отдельных признаков, определяется индивидуальными особенностями генотипа и наследуется. Важно знать, что модификации, идущие в пределах нормы реакции, имеют адаптивный (приспособительный) характер. Они обуславливают приспособление организмов к изменениям среды. В этом роль и значение фенотипической изменчивости для существования организмов.
Примеров модификационной изменчивости в природных популяциях много. Один из них - вариабельность формы листа у стрелолиста (Sagittaria sagittifolia). Это растение может расти на берегу реки или озера, или быть частично погруженным в воду. Встречаются отдельные экземпляры, целиком погруженные в воду. Надводные листья растения имеют стреловидную, плавающие по поверхности воды сердцевидную, подводные листья - характерную для водных растений лентовидную форму (рис. 1).
1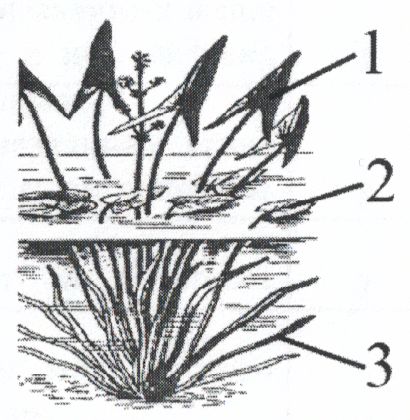 – воздушные;
– воздушные;
2 – надводные плавающие;
3 – водные, длинные узкие.
Рис. 1. Вариабельность формы листа стрелолиста
Одним из классических примеров модификационной изменчивости у животных являются опыты с изменением окраски шерсти у горностаевых кроликов. У этой породы мех белый, но отдельные части тела: лапы, уши, кончик морды, хвост - черные. Если на спине кролика, которая покрыта белой шерстью, выбрить небольшой участок и поместить животное в условия пониженной температуры, то на оголенном месте вырастет черная шерсть. Этот факт наглядно свидетельствует о роли одного из физических факторов среды - температуры в формировании такого признака, как окраска шерсти кролика. Если от таких кроликов получить потомство и выращивать в обычных условиях, то потомки будут иметь исходный тип окраски. Это доказывает, что данная изменчивость не затрагивает генотип, не передается по наследству и ограничивается изменением только фенотипа.
Что касается механизма фенотипической изменчивости, то следует признать, что механизмы ненаследственной изменчивости исследованы пока намного хуже механизмов наследственной изменчивости. Во многом эти механизмы, по-видимому, связаны с изменением экспрессии генов. Факторы среды могут изменять активность генов, действуя через различные регуляторные механизмы, что изменяет их фенотипическое проявление. Однако полностью сводить фенотипическую изменчивость к процессам регуляции активности гена, вероятно, нельзя. Существует явление спонтанной или случайной фенотипической изменчивости. Сущность случайной изменчивости состоит в том, что любому признаку в ходе его реализации при развитии организма свойственна известная степень чисто случайных колебаний, даже при идеальном постоянстве генотипа и среды. Считается, что подобная изменчивость присутствует всегда и отражает неустойчивость процессов фенотипической реализации генотипа.
В настоящее время широко ведутся работы по выявлению молекулярных механизмов фенотипической изменчивости. Это важно и для медицинской практики, так как знание возможности возникновения фенотипической изменчивости и механизмов ее развития позволяет оценить роль факторов внешней среды в развитии того или иного фенотипа и, следовательно, в развитии того или иного заболевания.
Многие модификации, особенно те, которые индуцируются физическими, химическими и другими факторами, резко изменяют фенотип особи, вызывая иногда уродства. Они не носят адаптивного характера и получили название морфозы. Иногда трудно отличить модификационные изменения, которые вызваны факторами среды, от изменений генетической природы в силу их сходства. Такие модификации получили название - фенокопии. Например, у женщин, перенесших в ранний срок беременности краснуху, нередко рождаются дети с врожденной катарактой (помутнение хрусталика глаза), которая не отличается от наследственной катаракты. Это надо понимать и при диагностике разграничивать заболевания генетической и фенотипической природы.
Генотипическая изменчивость
Генотипическая изменчивость связана с изменением генотипа. Приобретаемые вследствие этой изменчивости признаки передаются по наследству. Существует две формы генотипической изменчивости: комбинативная и мутационная.
Комбинативная изменчивость заключается в появлении новых признаков и свойств организма в результате образования иных комбинаций генов родителей в генотипах потомков. Комбинативная изменчивость связана с тремя явлениями: 1) с перекомбинацией генов при кроссинговере в профазу мейоза I, что обеспечивает образование новых комбинаций отцовских и материнских генов в хромосомах будущих гамет; 2) с независимым распределением хромосом при их расхождении в анафазу мейоза I, что сопровождается случайным комбинированием отцовских и материнских хромосом в будущих гаметах; 3) со случайным и равновероятным сочетанием гамет при оплодотворении. При оплодотворении в зиготе возникают новые комбинации генов и формируется генотип особи.
В процессе комбинативной изменчивости сами гены не меняются, меняется лишь сочетание их, что приводит к появлению потомков с новыми генотипами. Комбинативная изменчивость обеспечивает генетическое разнообразие особей в пределах вида и играет важную роль в эволюции.
Мутационная изменчивость
Термин «мутация» впервые был предложен Г. Де Фризом в его классическом труде «Мутационная теория» (1901). Мутацией Г. Де Фриз назвал явление скачкообразного, прерывистого изменения наследственного признака. В настоящее время термином «мутация» обозначают внезапные, наследуемые изменения генетического материала, которые могут возникать без видимых причин (спонтанно), либо быть индуцированы внешним воздействием на организм. Процесс возникновения мутаций именуют мутагенезом. Организм, приобретший какой-либо новый признак и тем самым изменивший свой фенотип в результате мутации, называют мутантом.
Теория мутаций была обоснована и сформулирована в 1901 г. Г. Де Фризом. Ее основные положения:
- мутации возникают внезапно без видимых переходов;
- возникшие новые формы в результате мутации вполне устойчивы;
- мутации являются качественными изменениями;
- мутации происходят в разных направлениях, они могут быть как полезными, так и вредными;
- одни и те же мутации могут возникать повторно.
По происхождению все мутации можно разделить на две группы: спонтанные и индуцированные. Спонтанные мутации происходят в естественных условиях без всякого видимого внешнего воздействия. Индуцированные мутации возникают под влиянием специальных воздействий (ионизирующей радиации, химических веществ, высокой температуры и др). Однако по механизму развития принципиальных различий между спонтанными и индуцированными мутациями нет.
По месту возникновения различают генеративные (герминальные) мутации, возникающие в половых клетках и передающиеся по наследству, и соматические, образующиеся в клетках тела, не участвующих в размножении. Соматические мутации часто ведут к появлению генетических мозаик, у которых измененной окажется лишь часть организма, развивающаяся в результате деления мутантных клеток.
По характеру изменения фенотипа различают: морфологические (видимые), физиологические и биохимические мутации.
Морфологические мутации проявляются изменением в строении организмов. К ним относятся, например, коротконогость у ряда сельскохозяйственных животных (овец, коров и т.д.), бескрылость у насекомых, гигантизм, карликовость, альбинизм у человека.
К физиологическим относят мутации, влияющие на жизнеспособность организмов, их развитие, ведущие к нарушению таких процессов как кровообращение, дыхание, умственная деятельность у человека, поведенческие реакции у животных и т.д.
Биохимические мутации представляют собой обширную группу, объединяющую все случаи изменения качества или количества, синтезируемых определенных химических веществ в организме, например, ферментов. Сюда относят различные мутации, нарушающие синтез ферментов, участвующих в репликации ДНК, репарации ее повреждений, транскрипции и трансляции генетического материала.
Классификация мутаций по фенотипу, по их проявлению условна. В действительности в основе проявления всех мутаций, как правило, лежат изменения биохимических процессов.
В зависимости от влияния на жизнеспособность и плодовитость мутации можно разделить на летальные, полулетальные, условно летальные, стерильные, нейтральные и усиливающие. К летальным относят мутации, ведущие к гибели зародышей, плода, что связано с отсутствием или недоразвитием жизненно важных органов у животных и человека.
Полулетальные мутации резко понижают жизнеспособность организмов и мутантные особи, как правило, не доживают до репродуктивного периода, быстро погибая из-за плохо совместимого с жизнью наследственного дефекта. Условно летальные мутации могут вовсе не проявляться в одних условиях и вести организм к гибели в других условиях. К стерильным относят мутации, существенно не затрагивающие жизнеспособность, но резко уменьшающие фертильность, или плодовитость, мутантов.
Наконец, к нейтральным и усиливающим относят мутации, не связанные с жизнеспособностью и плодовитостью, либо усиливающие эти признаки.
По характеру изменения наследственного материала мутации делят на: генные, хромосомные и геномные.
Генные мутации
Генные или точечные мутации - это стойкие изменения в молекулярной структуре отдельных генов. По молекулярным механизмам генные мутации подразделяют на два основных типа: мутации, вызванные заменой одного нуклеотида на другой, и мутации со сдвигом рамки считывания. Нуклеотиды, из которых состоят молекулы ДНК, различаются только по азотистому основанию, что дает право рассматривать главное событие генных мутаций первого типа как замену одного азотистого основания на другое. На их долю приходится порядка 20% спонтанных генных мутаций. Если в результате мутации пуриновые основания заменяются пуриновыми, а пиримидиновые - пиримидиновыми, то эти замены называют транзициями. Возможны четыре вида транзиций: А↔Г, Т↔Ц. Замены пиримидиновых азотистых оснований на пуриновые и, наоборот, называют трансверсиями. Трансверсии могут быть восьми видов: А↔Т, А↔Ц, Г↔Ц, Г↔Т. У человека транзиции встречаются чаще чем трансверсии.
Замены одного азотистого основания в макромолекуле ДНК приводит к образованию нового триплета в нуклеотидной последовательности, кодирующей последовательность аминокислот в полипептиде. Мутации замены оснований приводят к появлению двух типов триплетов в ДНК и соответственно кодонов в и(м)РНК - с измененным смыслом (миссенс-мутации) и бессмысленного (нонсенс- мутации). Результатом миссенс-мутаций, ведущих к изменению кодирующих триплетов, может быть замена одной аминокислоты в полипептиде белка на другую, что может иметь серьезные последствия для организма.
Примером мутаций такого типа может служить появление гемоглобина S при серповидно-клеточной анемии у человека. Суть такой миссенс-мутации сводится к замене второго нуклеотида (Т) в триплете, кодирующем стоящую в β-цепи гемоглобина на 6-м месте глутаминовую кислоту (ЦТТ), на нуклеотид (А), превращающий его в триплет, кодирующий аминокислоту валин (ЦАТ). Замена только одной аминокислоты в β-цепи гемоглобина приводит к тому, что вместо нормального гемоглобина А, состоящего из двух идентичных α-цепей и двух идентичных β-цепей, синтезируется гемоглобин S.
Необходимо отметить, что в силу вырожденности генетического кода в 25% таких замен возникает триплет-синоним, что не дает изменений аминокислотной последовательности в соответствующем полипептиде. Таким образом, не всякая мутация в триплете приводит к замене аминокислоты. Такие мутации называют нейтральными.
К типу нонсенс-мутаций относят мутации, приводящие к замене оснований в триплетах ДНК, при которой кодон и(м)РНК, определяющий какую-либо аминокислоту, превращается в один из терминирующих триплетов (стоп-кодонов), не транслирующихся на рибосомах. Появление такого триплета внутри структурного гена приводит к преждевременной терминации транскрипции и обрыву полипептидной цепи. Например, кодон УАЦ на и(м)РНК кодирует аминокислоту тирозин. При замене цитозина на аденин возникает бессмысленный кодон (УАА), который обрывает синтез полипептидной цепи. Говоря об однонуклеотидных заменах необходимо отметить, что однонуклеотидные замены наиболее частые патогенные мутации в геноме человека. Большинство этих мутаций возникает в ходе репликации ДНК, точного, но подверженного ошибкам процесса.
Немалую часть спонтанных (самопроизвольных) мутаций составляют мутации по типу сдвига рамки считывания. Мутации со сдвигом рамки обусловлены встраиванием в нуклеотидную последовательность ДНК дополнительных нуклеотидов (нуклеотидных пар) - инсерция, или выпадение нуклеотидов (нуклеотидных пар) - делеция. При этом меняются все триплеты после сайта (места) инсерции и делеции, что приводит к сдвигу рамки считывания информации и цепь полипептида приобретает цепь неверных аминокислот.
Инсерции и делеции могут представлять собой и нонсенс-мутации, поскольку вызывают появление терминирующих кодонов (УАА, УАГ, УГА), поэтому последствия таких мутаций довольно серьезны.
К выпадению или вставкам нуклеотидов (пар нуклеотидов) приводят достаточно частые мутации в сайтах сплайсинга - на участках интронов и экзонов. Эти мутации проявляются в нарушении вырезания интронов из пре-и(м)РНК транскрипта. В результате созревающая и(м)РНК лишается части или всего экзона, а может привести к сохранению в и(м)РНК интронной нуклеотидной последовательности.
Недавно стали выделять еще одну группу генных мутаций по типу экспансии тринуклеотидных повторов. Они характеризуются нарастанием числа триплетных повторов в кодирующих или некодирующих участках гена (например, ЦТТ, ЦГГ или ЦАГ). Это приводит к появлению в молекуле полипептида «трека» из аминокислотных остатков какой-либо аминокислоты.
Надо иметь ввиду, что всякая генная мутация есть результат сложного многоэтапного процесса, инициированного появлением в ДНК того или иного первичного повреждения. С этой точки зрения первичное повреждение, происходящие в какой-либо нити молекулы ДНК, являются всего лишь первым этапом в процессе образования истинной мутации (в процессе фиксации мутации). Такими типами первичных повреждений (предмутационных повреждений) в ДНК служат следующие:
1) химические изменения азотистых оснований (дезаминирование, окисление, алкилирование);
2) потеря основания (депуринизация, депиримидинизация);
3) димеризация пиримидинов (образованием димеров);
4) однонитевые и двунитевые разрывы ДНК;
5) ошибка спаривания.
Эти повреждения возникают в естественных условиях (спонтанные повреждения) и при воздействии мутагенных факторов на хромосомы (индуцированные). Спонтанные повреждения возникают без каких-либо направленных воздействий, а индуцированные – под действием физических, химических или биологических факторов. Причем частота спонтанных повреждений очень высокая. Ежедневно у человека возникает 50 тысяч однонитевых разрывов, более 8 тысяч окисленных и алкилированных оснований, и еще в совокупности около 100 сложных повреждений (двунитевые разрывы, межмолекулярные сшивки ДНК-ДНК и ДНК-белок). Ежедневно в каждой клетке человека от 2 до 3 тысяч пуриновых и пиримидиновы нуклеотидов (на гаплоидный геном) теряют свои азотистые основания. В результате образуются АП-сайты (апуриновые и апиримидиновые) и сохраняется только дезоксирибоза и фосфодиэфирная связь.
Первичные повреждения могут происходить в разных фазах клеточного цикла. Молекула ДНК может изменяться во время ее синтеза. Это касается, например, ошибок спаривания, когда во время синтеза новой нити молекулы ДНК при репликации вставляется неверное основание, или аналог его. Могут возникать повреждения и в покоящейся молекуле ДНК в фазе G1 и G2, вызывая повреждения нитей ДНК.
Итак, повреждения в ДНК происходят до, после или в процессе репликации первоначально, как правило, в одной полинуклеотидной цепи. Появление таких первичных изменений еще не означает появление мутаций. Из общего числа первичных повреждений только часть их переходит в истинные мутации (фиксируется в мутации). Остальные устраняются механизмами молекулярной репарации (см. ниже). Результатом превращения первичных повреждений в истинные генные мутации (результатом фиксации мутаций) служит: появление замен пар оснований (пар нуклеотидов), вставка (инсерция) или делеция пар оснований (пар нуклеотидов), их перестановки (рис. 1).
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Рис. 1. Основные типы генных мутаций 1 - исходный порядок пар нуклеотидов в гене; 2 - замена пар оснований ГЦ на АТ; 3 - вставка дополнительной пары оснований ТА; 4 - делеция пары оснований ТА.
|
||||||||||||||||
|
А |
Т |
Т |
Г |
Ц |
А |
Т |
Ц |
А |
|
|
||||||||||||||||||||||||||
1. |
II |
II |
II |
III |
III |
II |
II |
III |
II |
|
|
|
|||||||||||||||||||||||||
|
Т |
А |
А |
Ц |
Г |
Т |
А |
Г |
Т |
|
|
|
|||||||||||||||||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|||||||||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|||||||||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|||||||||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|||||||||||||||||
|
А |
Т |
Т |
Г |
Т |
А |
Т |
Ц |
А |
|
|
|
|||||||||||||||||||||||||
2. |
II |
II |
II |
III |
II |
II |
II |
III |
II |
|
|
|
|||||||||||||||||||||||||
|
Т |
А |
А |
Ц |
А |
Т |
А |
Г |
Т |
|
|
|
|||||||||||||||||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|||||||||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|||||||||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|||||||||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|||||||||||||||||
|
А |
Т |
Т |
Г |
Ц |
А |
Т |
Т |
Ц |
А |
|
||||||||||||||||||||||||||
3. |
II |
II |
II |
III |
III |
II |
II |
II |
III |
II |
|
||||||||||||||||||||||||||
|
Т |
А |
А |
Ц |
Г |
Т |
А |
А |
Г |
Т |
|
||||||||||||||||||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|||||||||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|||||||||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|||||||||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|||||||||||||||||
|
А |
Т |
Т |
Г |
Ц |
А |
Ц |
А |
|
|
|
|
|
||||||||||||||||||||||||
4. |
II |
II |
II |
III |
III |
II |
III |
II |
|
|
|
|
|
||||||||||||||||||||||||
|
Т |
А |
А |
Ц |
Г |
Т |
Г |
Т |
|
|
|
|
|
||||||||||||||||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|||||||||||||||||
В результате таких точечных мутаций искажается генетическая информация, она реализуется в неправильную последовательность аминокислот в соответствующих белках и ведет либо к появлению новых, отличающихся от нормы признаков (неоморфные мутации), либо к потере признака (аморфные мутации).
За появление спонтанных генных мутаций ответственны разные механизмы. Из них наиболее значимыми являются: ошибки репликации ДНК; неправильное спаривание оснований при кроссинговере; перемещение мобильных генетических элементов; нарушение процессов сплайсинга; генотоксическое действие продуктов метаболизма, повреждающих ДНК (активные формы кислорода и др.). Для естественного мутационного процесса имеет значение и уровень естественного фона радиации, который складывается из космических лучей, радиоактивных элементов поверхности Земли и радионуклидов, поступающих в организм.
Исследование спонтанных мутаций у различных организмов выявило некоторые закономерности. Во-первых, скорость спонтанного мутирования у разных организмов различна. Во-вторых, частота спонтанных мутаций в разных генах у организмов одного и того же вида также различна. У дрозофилы средняя частота мутаций на ген равна 1х10-5. Это значит одна новая мутация появляется на 100 тысяч гамет. У человека большинство генов мутирует с частотой от 1х10-5 до 1х10-6. Например, ген гемофилии мутирует с частотой 3х10-5. Это означает 3 новых мутации на 100 тысяч гамет. Однако обнаружены гены, которые в естественных условиях мутируют часто. Такие гены получили название высоко мутабельных (1х10-4 изменений на локус, на поколение). Для наиболее устойчивых участков генома человека частота мутаций 1х10-11. Однако следует отметить, что в целом спонтанные генные мутации представляют собой редкие случайные события.
Генные мутации - это обратимый процесс. Все дело в том, что нормальные особенности организмов вида, созданные в процессе эволюции, как правило, определяются доминантными генами (аллелями), наиболее распространенными в природе. Они получили название нормальных аллелей или генов «дикого типа». Мутационный процесс, идущий в нормальных особях, превращает доминантные нормальные аллели (гены «дикого типа») в мутантные - рецессивные. Изменения нормальных аллелей (генов «дикого типа») в мутантные называют прямыми мутациями (А→а), превращение мутантных рецессивных аллелей в нормальные доминантные называют обратными мутациями (а→А). Частота мутирования в прямом направлении (А→а), как правило, выше, чем частота обратных мутаций (а→А).
Как отмечалось ранее, мутации могут быть в соматических и половых клетках. Появление соматических мутаций может оказать большое влияние на жизнь особи. У взрослого человека число соматических клеток равно примерно 1014. Если мы примем, что ген мутирует с невысокой частотой 1х10-8, то в этом случае в организме человека должно содержаться более миллиона мутантных клеток только по данному гену. Если считать, что у человека около 25 тысяч генов, кодирующих белки, то огромная популяция клеток тела человека только по этим генам испытывает на себе постоянное действие мутаций.
Основной категорией мутаций следует признать мутации в половых клетках (герминальные мутации), поскольку они переходят в следующие поколения организмов и вступают в сложные рекомбинации в условиях полового размножения. У видов, размножающихся половым путем, потомству передаются лишь мутации, которые появляются в половых клетках. Вновь возникший от мутаций рецессивный аллель передается в гетерозиготной форме на протяжении многих поколений, пока скрещивание таких гетерозигот не даст гомозиготное потомство. У такого потомка мутантный аллель будет проявляться в фенотипе.
Мутационный процесс имеет место у всех организмов. Много мутаций обнаружено у дрозофилы. Например, желтое тело - рецессивная мутация, аллель расположен в Х-хромосоме; бескрылая форма - рецессивная мутация, мутантный аллель находится во II хромосоме. Большое количество генных мутаций открыто у человека. Они касаются его морфологии, биохимии, склонностей поведения, умственных способностей и т.д. Большинство наследственных заболеваний и другие отклонения от нормы в конечном счете - следствие генных мутаций.