

1.1.3. Полисахариды
Молекулы полисахаридов включают десятки, сотни и даже тысячи моносахаридных остатков, соединенных такими же гликозидными связя-ми, как и в составе олигосахаридов. Большинство из них образуют линей-ные полимеры, формирующие определенную пространственную структу-ру, однако некоторые полисахариды имеют разветвлённые молекулы. Мо-носахаридные остатки в составе полисахаридов находятся в циклической форме в виде а- или b-стереоизомеров.
Большинство полисахаридов представляют собой сложные углеводы, построенные из многократно повторяющихся остатков одного моносахари-да. Однако известны полисахариды, молекулы которых состоят из остат-ков разных моносахаридов.
По выполняемым функциям различают запасные и структурные полисахариды. Первые откладываются в клетках листьев или запасающих тканей в виде упорядоченных структур – гранул, вторые участвуют в по-строении клеточных стенок растений.
Крахмал. Крахмал – основное запасное вещество растений, пред-ставляющее собой смесь двух полисахаридов: амилозы и амилопектuна, различающихся по строению молекулы и физико-химическим свойствам. Однако молекулы этих полисахаридов построены из остатков одного моносахарида – а-D-глюкозы, находящейся в пиранозной форме.
В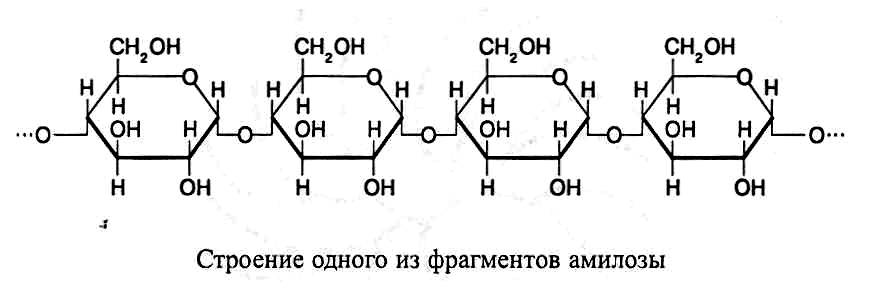 молекулах амилозы остатки a-D-глюкозы
соединены а(1®4)-свя-зями,
образуя
спиралевидно закрученные цепочечные
структуры, включа-ющие от 100 до 1–2 тыс.
глюкозных остатков. Молекулярная масса
амило-зы обычно составляет от 20 до 500
тыс. Спиралевидное закручивание
молекулы происходит вследствие
образования водородных связей между
остатками глюкозы, находящимися в
соседних витках. В каждом витке амилозы
содержится шесть пиранозных cтpyктyp,
соединенных в цепочку О-гликозидными
связями. Далее представлена схема
строения одного из фрагментов амилозы:
молекулах амилозы остатки a-D-глюкозы
соединены а(1®4)-свя-зями,
образуя
спиралевидно закрученные цепочечные
структуры, включа-ющие от 100 до 1–2 тыс.
глюкозных остатков. Молекулярная масса
амило-зы обычно составляет от 20 до 500
тыс. Спиралевидное закручивание
молекулы происходит вследствие
образования водородных связей между
остатками глюкозы, находящимися в
соседних витках. В каждом витке амилозы
содержится шесть пиранозных cтpyктyp,
соединенных в цепочку О-гликозидными
связями. Далее представлена схема
строения одного из фрагментов амилозы:
Амилоза растворяется в теплой воде и при добавлении раствора йода в водном растворе йодида калия окрашивается в синий цвет, поскольку йод образует комплeксы с остатками глюкозы. Водные растворы амилозы не отличаются высокой вязкостью и при стоянии довольно быстро обра-зуют кристаллический осадок.
Амилопектин имеет разветвленные молекулы, построенные из остат-ков а-D-глюкозы. В точках ветвления гликозидные связи образуются меж-ду первым и шестым углеродными атомами глюкозных остатков – а(I®6)-связи. Между точками ветвления глюкозные остатки так же, как в амилозе,
с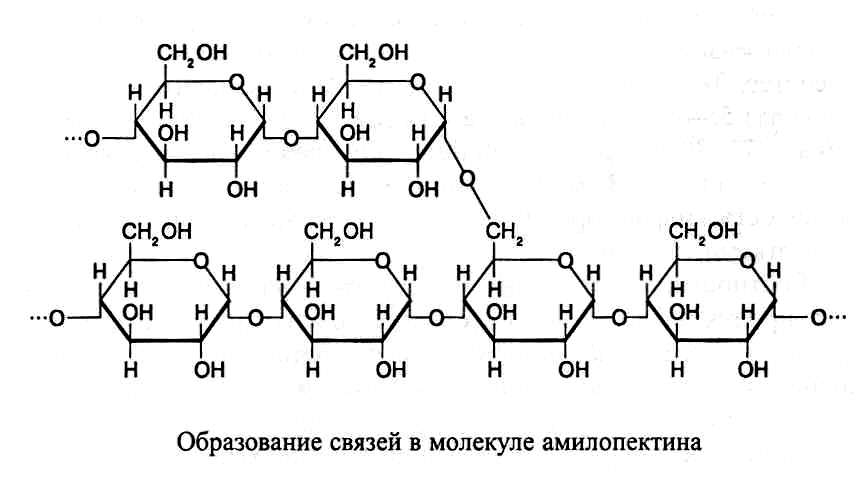 вязи.
Между точками ветвления глюкозные
остатки так же, как в амилозе, соединены
а(I®4)-связями:
вязи.
Между точками ветвления глюкозные
остатки так же, как в амилозе, соединены
а(I®4)-связями:
Ответвления в молекулах амилопектина образуются через каждые 12–15 остатков глюкозы. Молекулярная масса амилопектина значительно больше, чем амилозы, и может достигать 1 млн. Схема строения молекулы амилопектина показана на рисунке 1.1.
А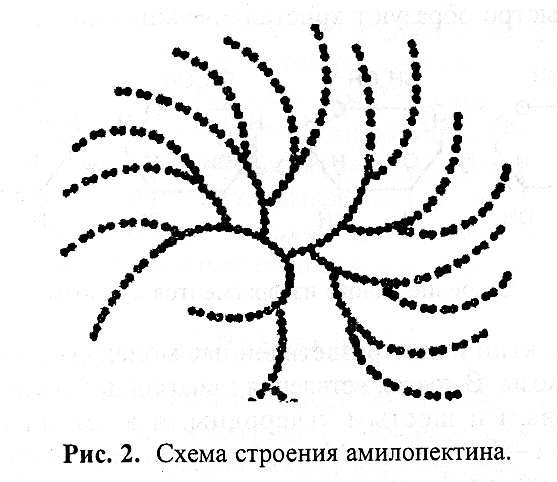 милопектин
в теплой воде не растворяется, а при
более сильном на-гревании с водой
образует очень вязкий коллоидный раствор
– клейстер. Температура клейстеризации
картофельного и ржаного крахмала –
55–65°С,
пшеничного и кукурузного – 60–70, крахмала
риса – 70–80 °C.
Йодным реактивом амилопектин окрашивается
в красно-фиолетовый цвет. В амилопектине
в небольшом количестве содержатся
остатки ортофосфор-ной
кислоты,
соединенные эфирной связью с остатками
глюкозы.
милопектин
в теплой воде не растворяется, а при
более сильном на-гревании с водой
образует очень вязкий коллоидный раствор
– клейстер. Температура клейстеризации
картофельного и ржаного крахмала –
55–65°С,
пшеничного и кукурузного – 60–70, крахмала
риса – 70–80 °C.
Йодным реактивом амилопектин окрашивается
в красно-фиолетовый цвет. В амилопектине
в небольшом количестве содержатся
остатки ортофосфор-ной
кислоты,
соединенные эфирной связью с остатками
глюкозы.
Рис. 1.1. Схема строения амилопектина
Соотношение амилозы и амилопектина в различных растительных продуктах варьирует в очень широких пределах. В картофельном крахмале на долю амилозы приходится около 20 %, пшеничном и кукурузном – око-ло 25, рисовом – 15–20, в крахмале гороха и некоторых сортов кукурузы – 50–80 %. Крахмал яблок почти полностью состоит из амилозы, а крахмал восковидных сортов кукурузы – только из амилопектина.
У одного и того же вида растений содержание амилозы и амилопектина в крахмале может изменяться в зависимости от фазы их разви-тия и условий внешней среды. В разных органах растений синтезируется крахмал совершенно определенного состава. Так, например, в крахмале клубней картофеля обычно содержится 19–22 % амилозы, а в молодых побегах – в два раза больше.
В растениях крахмал образуется в листьях как продукт фотосинтеза, а в семенах, клубнях, корневищах, утолщенных частях стеблей, древесине – как запасное вещество. Фотосинтетический крахмал откладывается в хлоропластах в виде гранул, называемых крахмальными зернами, и до-вольно быстро используется в процессе дыхания и для синтеза других веществ. Значительная его часть превращается в транспортную форму углеводов – сахарозу, которая поступает по флоэме в нефотосинтезирую-щие органы, распадается там до глюкозы и фруктозы и в виде этих моносахаридов включается в различные биохимические процессы.
Запасной крахмал также откладывается в виде крахмальных зерен и у целого ряда растений накапливается в значительном количестве в запасающих тканях и органах. В зерне злаковых культур его содержание обычно составляет 40–70 % (в рисе – до 80), зерне зернобобовых культур – 35–50; клубнях картофеля – 11–20, батата, ямса и маниока – 20–30; в корнеплодах моркови – до 1 %; листьях растений (включая листовые овощи) – до 1–2 %; в семенах лиственных древесных растений – до 50–60 %, хвойных пород – до 20, древесине – 3–10.
Крахмальные зерна чаще всего имеют вид овальных или сферических частиц (рис. 1.2), имеющих разную форму и размеры (2–170 мкм). Под микроскопом можно различить слоистое строение крахмальных зерен. Их размеры и строение у разных видов и даже сортов растений имеют харак-терную специфику, что можно использовать для идентификации геноти-пов, а также обнаружения примесей одного растительного продукта в другом.
Запасной крахмал вначале откладывается в пластидах, называемых амилопластами. По мере наполнения происходит постепенная деградация их мембранной структуры и они превращаются в крахмальные зерна.
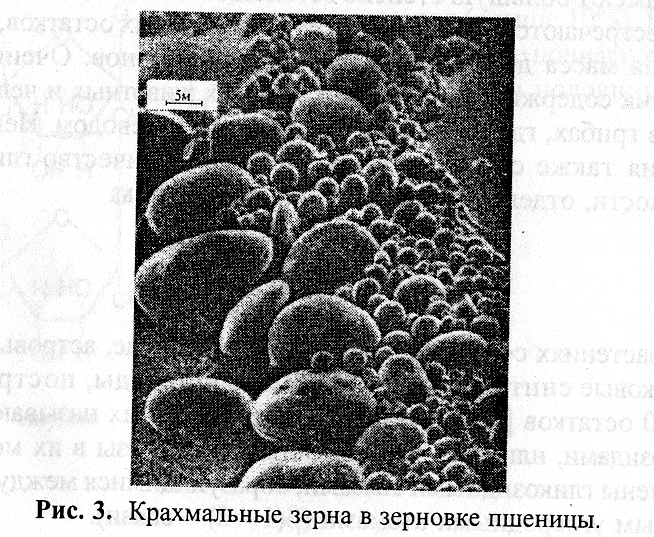 Рис. 1.2.
Крахмальные зерна в зерновке пшеницы
Рис. 1.2.
Крахмальные зерна в зерновке пшеницы
Крахмалоносные растения представляют собой легковозобновляемое сырье для перерабатывающей промышленности, которое используют для получения продовольственного и технического крахмала, глюкозы, этило-вого спирта и даже пластмасс, обладающих высокой прочностью и эколо-гической безопасностью (при сгорании не образуют ядовитых веществ).
Гликоген. Гликоген – структурный аналог амилопектина. Его моле-кулы построены из остатков a-D-глюкозы, но по сравнению с амилопектином имеют большую степень ветвления. Точки ветвления у гликогена образуются через каждые 8–10 глюкозных остатков, а молекулярная масса достигает нескольких миллионов. Очень много гликогена содержится в печени и мышцах животных и человека, а также в грибах, где он является резервным углеводом. Некоторые растения, например, отдельные разно-видности кукурузы, также синтезируют небольшое количество гликогена.
Полифруктозиды. В растениях семейств Лилейные, Мятликовые, Астровые, Колокольчиковые синтезируются запасные углеводы, построен-ные из остатков b-D-фруктозы, в связи с чем их называют полифрукто-зидами или фруктанами. Полифрутозиды содержатся в листьях, корнях, семенах этих растений, накапливаются в значительном количестве в нижней утолщенной части стеблей мятликовых трав (до 6–8 % сухой массы) и в созревающих зерновках злаковых культур (рожь, пшеница, ячмень, овес). В листьях они являются основными продуктами фотосинте-за, тогда как фотосинтетический крахмал у этих растений не образуется.
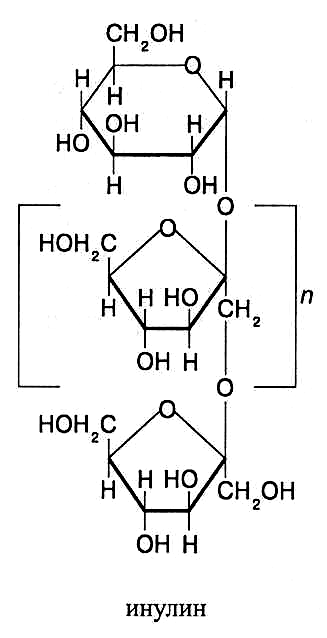
строение молекул полифруктозидов инулинового типа
Из полифруктозидов наиболее хорошо изучен инулин, содержащий в молекуле 37–44 фруктозных остатка, которые соединены β (1→2)-связями. К одному из концов молекулы инулина присоединен остаток а-D-глюкозы. Молекулярная масса инулина 5–6 тыс. Он хорошо растворяется в горячей воде, не обладает восстановительными свойствами. Растения, способные накапливать инулин, используют как кормовые культуры и сырье для про-мышленного производства фруктозы, которую получают из инулина путём его кислотного гидролиза.
Большое количество инулина содержится в клубнях георгина и арти-шока (до 50 %), топинамбура (10–12 %), корнях цикория (свыше 10 %). В чесноке общее содержание полифруктозидов достигает 20–30 % и поло-вину из них составляет инулин. Кроме инулина, в вегетативной массе и семенах злаковых растений содержатся другие полисахариды инулинового типа, содержащие в молекулах от 4 до 50 фруктозных остатков, которые соединены β (1→2)-связями и присоединены к фруктозному остатку моле-кулы сахарозы.
В вегетативной массе и семенах мятликовых трав, а также в клетках некоторых микроорганизмов (включая почвенные бактерии) синтезируют-ся полифруктозиды, называемые леванами, у которых остатки фруктозы соединены О-гликозидными связями, образующимися между шестым и вторым углеродными атомами – β (6→2)-связи. У этих полисахаридов полифруктозная цепь присоединена к фруктозному остатку молекулы сахарозы и она может содержать до 200 остатков фруктозы.
В зерновках ржи, пшеницы и ячменя содержатся полифруктозиды грамининового типа (граминин – полифруктозид ржи), включающие от 4 до 10 фруктозных остатков, которые присоединены к фруктозному остатку молекулы сахарозы β (1→2)-связью и к ее глюкозному остатку – β (6→2)-связью. В полифруктозной цепи остатки фруктозы соединены β (1→2)-связями.
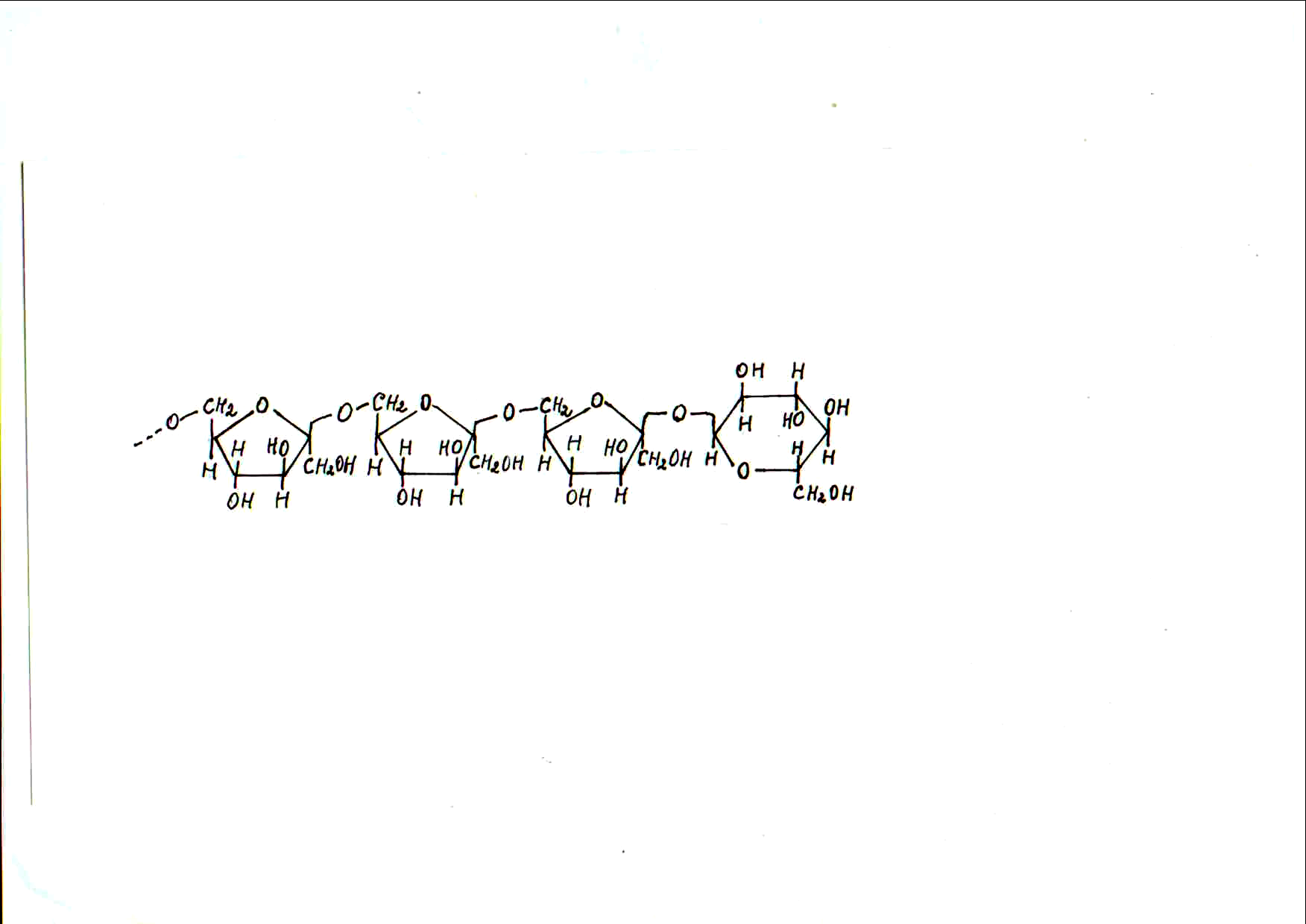
Ц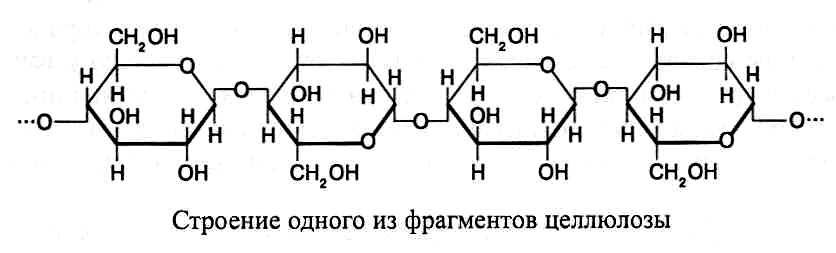 еллюлоза.
Целлюлоза
(клетчатка) – довольно устойчивое
вещество волокнистого строения, не
растворяется в воде и органических
раствори-телях, однако хорошо растворима
в аммиачном растворе гидроксида меди
(реактив Швейцера). Молекулы целлюлозы
состоят из остатков b-D-глюкозы
в пиранозной форме,
соединенных
b
(1®4)-связями.
В каждой молекуле целлюлозы может
содержаться 1,5–10 тыс. пиранозных остатков
b-D-глюкозы,
образующих неразветвленный полимер.
Далее представлена схема строения
одного из фрагментов молекулы целлюлозы:
еллюлоза.
Целлюлоза
(клетчатка) – довольно устойчивое
вещество волокнистого строения, не
растворяется в воде и органических
раствори-телях, однако хорошо растворима
в аммиачном растворе гидроксида меди
(реактив Швейцера). Молекулы целлюлозы
состоят из остатков b-D-глюкозы
в пиранозной форме,
соединенных
b
(1®4)-связями.
В каждой молекуле целлюлозы может
содержаться 1,5–10 тыс. пиранозных остатков
b-D-глюкозы,
образующих неразветвленный полимер.
Далее представлена схема строения
одного из фрагментов молекулы целлюлозы:
Между линейно вытянутыми молекулами целлюлозы, имеющими свободные гидроксильные группы, возникают водородные связи, с по-мощью которых нитевидные полимеры, построенные из остатков глюкозы, объединяются в пучки, включающие несколько десятков молекул. Такие целлюлозные пучки, или фибриллы, обладают очень высокой прочностью и служат структурной основой клеточных стенок растений. Как видно на электронной микрофотографии клеточной оболочки (рис. 1.3), целлюлоз-ные фибриллы размещаются слоями, образуя сетчатую структуру, сквозь которую свободно проникает жидкая физиологическая среда с растворен-ными в ней веществами.
Ц еллюлоза
в том или ином количестве содержится
во всех тканях растений. Особенно много
целлюлозы в растительных волокнах
(хлопко-вом, льняном) – 80–95 %, древесине
и соломе – 40–50 %. В других рас-тительных
продуктах ее значительно меньше, %: в
зерне злаковых и зернобобовых культур
– 2–6, семенах пленчатых злаков – 7–14,
семенах масличных культур – 5–25, клубнях
картофеля – около 1, корнеплодах –
0,5–1,5, овощах 0,5–1,2 (томатах – 0,2,
капусте – 1–2), плодах и ягодах – 0,5–2,
вегетативной массе кормовых трав –
20–35, в молодых побегах сосны (вместе с
хвоей) – 20–25 (последние два показателя
даны в расчете
на сухую массу).
еллюлоза
в том или ином количестве содержится
во всех тканях растений. Особенно много
целлюлозы в растительных волокнах
(хлопко-вом, льняном) – 80–95 %, древесине
и соломе – 40–50 %. В других рас-тительных
продуктах ее значительно меньше, %: в
зерне злаковых и зернобобовых культур
– 2–6, семенах пленчатых злаков – 7–14,
семенах масличных культур – 5–25, клубнях
картофеля – около 1, корнеплодах –
0,5–1,5, овощах 0,5–1,2 (томатах – 0,2,
капусте – 1–2), плодах и ягодах – 0,5–2,
вегетативной массе кормовых трав –
20–35, в молодых побегах сосны (вместе с
хвоей) – 20–25 (последние два показателя
даны в расчете
на сухую массу).
Рис. 1.3. Микрофибриллы целлюлозы в составе
клеточной стенки растения
Целлюлоза практически не усваивается организмами человека и не-жвачных животных, тогда как жвачные животные способны ее усваивать с помощью ферментов микроорганизмов, обитающих в преджелудках этих животных и участвующих в процессах пищеварения.
При нагревании с раствором кислоты целлюлоза подвергается гидро-лизу, превращаясь в глюкозу, которая используется как источник углерода для культивирования дрожжевых клеток с целью промышленного получе-ния этилового спирта и кормовых дрожжей с повышенным содержанием белков и витаминов. При этом в качестве источника целлюлозы служат отходы древесины и целлюлозосодержащие растительные остатки – соло-ма, корзинки подсолнечника, льняная костра, стержни кукурузных почат-ков, свекловичная меласса, картофельная мезга, хлопковая шелуха и др. Большое количество целлюлозы расходуется для химической переработки.
В построении клеточных стенок растений наряду с целлюлозой участвуют также другие структурные полисахариды – гемицеллюлозы и пектиновые вещества, которые связаны с молекулами целлюлозы водород-ными связями.
Гемицеллюлозы. Гемицеллюлозы – это смесь полисахаридов, обра-зующих при гидролизе маннозу, галактозу, ксилозу, арабинозу и уроновые кислоты (глюкуроновую и галактуроновую). Они нерастворимы в воде, но растворяются в щелочных растворах. В клеточных стенках растений содер-жание гемицеллюлоз составляет около 30 %. Много их накапливается в древесине и соломе (10–30 %), оболочках семян, кукурузных початках, отрубях, вегетативной массе растений; в молодых побегах сосны (вместе с хвоей) – 8–10 % сухой массы. Разные виды растений заметно различаются по составу гемицеллюлоз. В древесине лиственных пород деревьев много содержится пентозанов, в древесине хвойных – гексозанов.
М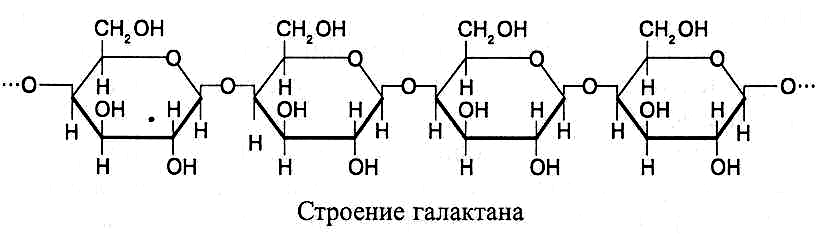 олекулы
галактанов
построены из остатков b-D-галактозы
в пиранозной форме, соединенных b
(1®4)-связями.
В каждой молекуле объединяются более
100 остатков галактозы. Строение галактанов
можно представить следующим образом:
олекулы
галактанов
построены из остатков b-D-галактозы
в пиранозной форме, соединенных b
(1®4)-связями.
В каждой молекуле объединяются более
100 остатков галактозы. Строение галактанов
можно представить следующим образом:
Галактаны входят в состав клеточных стенок растений, много их содержится в семенах люпина и некоторых древесных растений, а также древесине хвойных деревьев.
В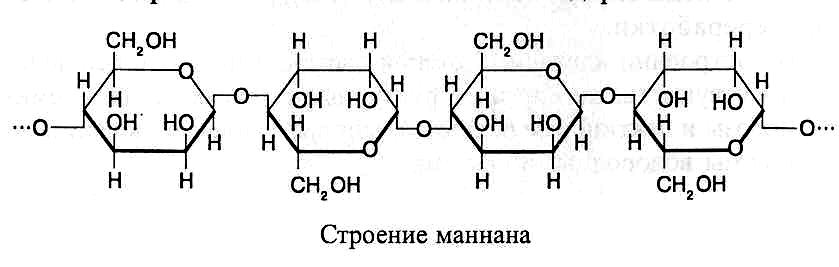 молекулах маннанов
остатки маннозы соединены b
(1®4)-связями.
В каждой молекуле насчитывается 200–400
моносахаридных остатков,
на-ходящихся
в пиранозной форме. Много маннанов
содержится в семенах некоторых лиственных
древесных растений и древесине хвойных
деревьев. Соединение остатков маннозы
в молекуле маннана можно показать в
виде следующей схемы:
молекулах маннанов
остатки маннозы соединены b
(1®4)-связями.
В каждой молекуле насчитывается 200–400
моносахаридных остатков,
на-ходящихся
в пиранозной форме. Много маннанов
содержится в семенах некоторых лиственных
древесных растений и древесине хвойных
деревьев. Соединение остатков маннозы
в молекуле маннана можно показать в
виде следующей схемы:
 В
состав молекул ксиланов
входят остатки b-D-ксилозы
в пираноз-ной форме, соединенные b
(1®4)-связями.
В структуре полимера могут находиться
до 200 ксилозных остатков. В соломе и
других растительных отходах содержание
ксиланов достигает 25–28 %, в древесине
лиственных деревьев – до 25, хвойных –
до 12 %.
В
состав молекул ксиланов
входят остатки b-D-ксилозы
в пираноз-ной форме, соединенные b
(1®4)-связями.
В структуре полимера могут находиться
до 200 ксилозных остатков. В соломе и
других растительных отходах содержание
ксиланов достигает 25–28 %, в древесине
лиственных деревьев – до 25, хвойных –
до 12 %.
строение фрагмента молекулы ксилана
В молекулах ксиланов обычно имеются ответвления в виде остатков арабинозы, а также глюкуроновой и галактуроновой кислот. Ответвления чаще всего образуются путем этерификации третьего углеродного атома ксилозы. Карбоксильные группы остатков уроновых кислот образуют эфи-ры с метиловым спиртом. Ксиланы разных растений отличаются частотой и моносахаридным набором ответвлений в молекуле.
Арабаны
– полисахариды клеточной стенки
растений, которые состо-ят из остатков
a-L-арабинозы,
соединенных гликозидной связью между
первым и пятым углеродными атомами. При
этом к каждому
второму ос-татку арабинозы в линейной
структуре присоединен в виде
ответвления еще один остаток арабинозы.
В ответвлениях связь образуется между
т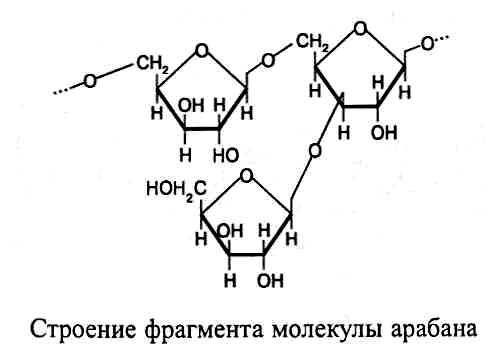 ретьим
углеродным атомом арабинозы, находящейся
в цепочке, и первым углеродным атомом
бокового остатка арабинозы:
ретьим
углеродным атомом арабинозы, находящейся
в цепочке, и первым углеродным атомом
бокового остатка арабинозы:
Как и целлюлоза, гемицеллюлозы не усваиваются организмом чело-века, но могут усваиваться жвачными животными с помощью ферментов микроорганизмов, находящихся в преджелудках.
К глюканам относятся полисахариды, образуемые из b-D-глюкозы, но в их молекулах остатки глюкозы соединяются не только b (1®4)-связями, как в целлюлозе, но также и b (1®3)-связями или только ими. К таким полисахаридам относятся каллоза и лихенин. Каллоза – полисаха-рид, молекула которого включает до 100 остатков b-D-глюкозы, соединен-ных b (1®3)-связями. Она содержится в ситовидных трубках флоэмной системы растений. В молекулах лихенина остатки b-D-глюкозы соединены как b (1®4)-, так и b (1®3)-связями (встречаются с частотой около 30 %). Лихенин входит в состав клеточных стенок растений, а также углеводного комплекса зерна овса и ячменя, особенно много его в лишайниках.
Пектиновые вещества. Пектиновые вещества в растениях представ-лены двумя группами соединений – пектинами и протопектинами, которые