

5.6. Особенности желудочного пищеварения у некоторых видов животных
Желудочное пищеварение у лошади. Желудок у лошадей однокамерный (рис. 5.5). В зависимости от размеров лошади его емкость колеблется от 6 до 15 л. В желудке различают кардиальную, фундальную и пилорическую части. Кардиальная часть имеет расширенный слепой мешок, и в ней отсутствуют железы. В слизистой оболочке фундальной и пилорической частей расположены кардиальные, фундальные и пилорические железы. В желудке рН содержимого колеблется от 1,1 до 6,8, а концентрация соляной кислоты — от 0,05 до 0,29 %. В слепом мешке благодаря поступлению слюны сохраняется слабощелочная реакция, что способствует жизнедеятельности микрофлоры. Уже в слепом мешке начинается переваривание клетчатки и крахмала, но в меньшей степени, чем в преджелудках жвачных. В процессе пищеварения также принимают участие ферменты самого корма. Растительные диастазы в слабощелочной среде расщепляют крахмал до дисахаридов и глюкозы, чему способствует характерное для желудка лошади послойное распределение корма. Желудочный сок постепенно пропитывает и разжижает пищевой ком, поэтому внутри его в течение некоторого времени поддерживается слабощелочная среда. Слюна также способствует поддержанию щелочной реакции содержимого желудка в его кардиальной и центральной частях.
В пищевой массе, уже пропитанной желудочным соком, начинают действовать его ферменты — пепсины и липаза. Таким образом, в желудке лошади происходит одновременное переваривание
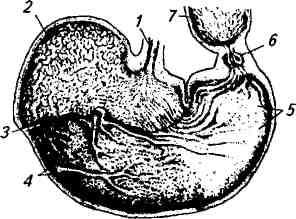
Рис. 5.5. Желудок лошади:
/ — кардиальный сфинктер; 2— пищеводная часть; 3 — граница между пищеводной и фундальной частью; •/—область дна; 5— пилори-ческая часть; 6— пилорический сфинктер; 7— двенадцатиперстная кишка
углеводов, белков и жиров. Переваривание в желудке и эвакуация пищевого кома в двенадцатиперстную кишку — медленно протекающие процессы.
Секреция желудочного сока у лошади происходит непрерывно, а каждый прием корма усиливает ее. Даже пустой желудок выделяет за сутки в среднем до 30 л сока. Эта особенность позволила использовать лошадей в качестве продуцентов желудочного сока. В связи с непрерывным характером желудочной секреции фазы секреторной деятельности желудка у лошади выражены слабо.
Моторная функция желудка характеризуется периодичностью и волнообразностью. При снижении кислотности деятельность ее усиливается. По мере переваривания и эвакуации пищи кислотность желудочного сока снижается, а моторная деятельность возрастает.
Желудочное пищеварение у свиней. По морфофункциональ-ным особенностям пищеварительных органов свиньи занимают промежуточное положение между травоядными животными с многокамерным желудком и плотоядными, у которых желудок однокамерный.
В однокамерном желудке свиньи (рис. 5.6) в отличие от желудка плотоядных животных кардиальная часть занимает около половины общей площади. В области впадения пищевода кардиальная часть желудка образует выпячивание — слепой мешок. По строению слизистую оболочку в желудке подразделяют на следующие зоны: пищеводную, кардиальную, дивертикул слепого мешка, зону дна желудка и пилорическую. В пищеводной зоне отсутствуют железы. В слизистой оболочке слепого мешка и кардиальной зоны расположены железы, вырабатывающие слизистый секрет, не содержащий пепсина и соляной кислоты. Железы фундальной и пилорической зон соответствуют таковым у плотоядных животных. Желудочный сок свиней содержит ферменты пепсин и химозин, наличие липазы точно не установлено.

Рис. 5.6. Желудок свиньи: 2
I — пилорический сфинктер; 2 — пилорическая
часть; 3 — дно желудка; 4 — пищеводная часть;
5— слепой мешок; 6— кардиальная часть
Желудочная секреция у свиней происходит непрерывно. В связи с актом приема пищи желудочная секреция резко усиливается. Реф-
206
207
 лекторная
фаза желудочной секреции наступает
через 5—10 мин от начала
кормления и продолжается около двух
часов.
лекторная
фаза желудочной секреции наступает
через 5—10 мин от начала
кормления и продолжается около двух
часов.
Особенностью желудочного пищеварения у свиней является слабое перемешивание пищевых масс при поедании кормов плотной консистенции и послойное распределение корма по мере его поедания. В желудке свиньи, так же как и лошади, одновременно идут амилолитические и протеолитические процессы. В кардиальной зоне желудка и слепом мешке создаются благоприятные условия для переваривания углеводов с помощью ферментов слюны и растительных кормов. В фундальной и пилорической частях под действием пепсина и соляной кислоты идет гидролиз белков до пептонов. В желудке свиньи происходит и молочнокислое брожение, но его интенсивность относительно невелика.
У новорожденных поросят желудочный сок не содержит свободной соляной кислоты. Кислая реакция в желудке обусловлена в этот период присутствием молочной кислоты, которая образуется при расщеплении гликогена, находящегося в клетках желудочных желез. Соляная кислота появляется в желудочном соке ко второй-третьей декаде.
Желудочное пищеварение у жвачных. Жвачные животные составляют достаточно крупную группу животных (крупный и мелкий рогатый скот, северные олени, верблюды и т. д.). Основными признаками, объединяющими животных этой группы, является наличие многокамерного желудка и жвачного процесса.
Желудок жвачных состоит из четырех отделов: рубец, сетка, книжка, сычуг (рис. 5.7). Из них только сычуг — истинный желудок, имеющий секреторные железы, продуцирующие желудочный сок. Рубец, сетка, книжка относятся к преджелудкам; пищеварение в них происходит за счет симбионтной микрофлоры рубца. В рубце переваривается значительная часть углеводов, липидов и белков корма.
В преджелудках происходит механическая,
химическая, биологическая
обработка корма, расщепление и синтез
питательных веществ. Пищевая
масса, поступающая в сычуг, по своему
составу существенно отличается
от съеденного корма.
преджелудках происходит механическая,
химическая, биологическая
обработка корма, расщепление и синтез
питательных веществ. Пищевая
масса, поступающая в сычуг, по своему
составу существенно отличается
от съеденного корма.
Рис. 5.7. Желудок жвачных:
1—рубец; 2— конечный участок пищевода; 3— пищеводный желоб; 4— сетка; 5— книжка; 6— сычуг
Пищеварительные функции рубца. Из пищевода смоченный слюной пищевой ком попадает в рубец. Емкость рубца у овец составляет 4...10 л, у крупного рогатого скота— 100...300 л. Рубец представляет собой большую бродильную камеру. Съеденный корм в рубце измельчается — этому способствует периодически повторяющаяся жвачка и только тогда переходит в последующие отделы пищеварительного тракта. В рубце переваривается до 50 % сухого вещества рациона. Характерной особенностью рубца является наличие богатой симбионтной микрофлоры, представленной более 100 видами бактерий и одноклеточных простейших (например, инфузории). Все симбионтные бактерии — анаэробы. Наиболее распространенные в рубце микроорганизмы — малые палочки и кокки (стрептококки, молочнокислые бактерии, целлюлолити-ческие бактерии). Нормальная жизнедеятельность рубцовой микрофлоры обусловлена постоянством среды их обитания в полости рубца за счет следующих факторов:
частое поглощение корма и задержка его в рубце до 7...8 сут обеспечивает регулярное поступление субстратов для микроорганизмов;
растворимые продукты жизнедеятельности микроорганизмов легко всасываются в кровь или переводятся в другие отделы желудка, не накапливаясь в рубце;
температура в рубце поддерживается в пределах 38...42 "С, причем ночью она выше, чем днем;
слюна жвачных богата бикарбонатом; за счет нее в основном поддерживается объем жидкости, постоянство рН и ионного состава. За сутки в рубец поступает около 300 г NaHC03. В ней содержится также значительное количество мочевины и аскорбиновой кислоты, имеющих важную роль для жизнедеятельности симбионтной микрофлоры.
В процессе переваривания корма в рубце образуются газы в результате биохимической активности микроорганизмов. За сутки может образовываться до 1000 л газов в зависимости от вида корма: диоксид углерода (до 60...70 %), метан (40...50 %), азот, ацетон, летучие жирные кислоты (ЛЖК) и небольшое количество водорода, сероводорода и кислорода. Избыток газов из рубца в основном удаляется при отрыжке. Наибольшее газообразование происходит при поедании зеленых сочных кормов, особенно бобовых.
Переваривание углеводов. Переваривание клетчатки в рубце происходит за счет целлюлолитической активности симбионтных микроорганизмов. Клетчатка (или целлюлоза) — основная составляющая кормов, входящих в рацион жвачных животных. В растительных кормах ее содержится до 40...50 %. Инфузории и другие простейшие, разрыхляя клетчатку, подготавливают ее для воздействия ферментов. Гидролиз целлюлозы осуществляется с помощью нескольких бактериальных ферментов, ответ-
208
14 — 3389
209
ственных за разные этапы расщепления ее макромолекул. Промежуточным продуктом расщепления клетчатки является дисахарид целлобиоза, который, в свою очередь, расщепляется до глюкозы. При этом фермент, отвечающий за начальный этап гидролиза, более лабилен и менее требователен к условиям среды, чем ферменты, катализирующие гидролиз уже частично расщепленного субстрата. Степень гидролиза клетчатки у жвачных снижается, если в корме увеличивается доля легкосбраживаемых углеводов (например, крахмала).
Крахмал в рубце может расщепляться до ди- и моносахаридов. В расщеплении крахмала принимают участие инфузории и бактерии. В рубце содержатся амилазы как растительного, так и микробного происхождения. Кроме того, некоторые простейшие (инфузории) способны захватывать и переваривать зерна крахмала, при этом большая часть субстрата накапливается в виде гликогена или амилопектина и затем сбраживается в течение длительного времени с почти постоянной скоростью. Это предупреждает внезапные вспышки брожения при поступлении в рубец свежего корма.
Простые сахара, поступившие с кормом или образовавшиеся в результате гидролиза макромолекул, активно используются как простейшими, так и бактериями рубца и сбраживаются до летучих жирных кислот. Брожению подвергаются и безазотистые остатки аминокислот. При сбраживании Сахаров образуются жирные кислоты — молочная, уксусная, пропионовая, масляная и жидкие нелетучие. В рубце чаще преобладает уксусная кислота, в небольших количествах встречаются изомеры масляной и валериановой кислот.
Таким образом, образующиеся в результате гидролиза сахара (в том числе глюкоза) не успевают всасываться в кровь до превращения в ЛЖК. Поэтому концентрация глюкозы в крови у жвачных намного ниже, чем у моногастричных животных. Однако поступившие в кровь Рубцовых вен ЛЖК замещают функции глюкозы: выступают в качестве источника энергии и участвуют в синтезе сложных углеводов и липидов. В частности, большое значение ЛЖК имеют в синтезе молочного жира. Введение в рационы лактирую-щих коров солей уксусной кислоты (например, ацетата натрия) значительно увеличивает удои и повышает жирность молока.
Таким образом, по сравнению с другими видами животных у жвачных значительно изменен углеводный обмен.
Переваривание белка. Белки, поступившие в рубец с кормом и слюной, расщепляются под действием протеолитичес-ких ферментов микроорганизмов до пептидов, аминокислот, а затем до аммиака. Пептиды, аминокислоты и аммиак используются бактериями для синтеза бактериального белка, имеющего для жвачных определенную питательную ценность. В процессе синтеза основная часть белка корма (40...80%) превращается в белок
микроорганизмов, а остальной белок в неизмененном виде поступает в сычуг и кишечник. В сутки у коров образуется до 100 г микробного белка, причем этот белок содержит, в отличие от белка растительного происхождения, все незаменимые аминокислоты, т. е. является биологически полноценным.
Микроорганизмы рубца могут использовать не только белок, но и небелковые азотистые вещества корма. Поэтому оказалось возможным часть белка в рационе жвачных животных заменить синтетической мочевиной (карбамидом). В рубце карбамид расщепляется ферментом уреазой, выделяемой микроорганизмами, до аммиака и диоксида углерода. В рацион, содержащий карбамид в качестве заменителя белка, должны входить корма, богатые легко-переваримыми углеводами, для нормальной жизнедеятельности организмов, использующих карбамид и аммиак. В противном случае карбамид не усваивается в рубце и всасывается в кровь в неизмененном виде, вызывая сильную интоксикацию.
Моторика преджелудков. Сокращения отдельных частей пред-желудков скоординированы между собой.
В норме рубец сокращается 2...5 раз в 2 мин. При этом происходит последовательное сокращение его отделов — преддверия рубца, дорсального мешка, вентрального мешка, каудодорсально-го слепого выступа, каудовентрального слепого выступа, а затем снова дорсального и вентрального мешков.
Сокращение дорсального мешка сопровождается отрыгивани-ем газов. При сокращении вентрального мешка плотное рубцовое содержимое отжимается в сторону левой голодной ямки, а разжиженная масса выжимается в сторону преддверия рубца. При этом поперечная складка опускается вниз и полужидкая масса заполняет преддверие рубца и сетку. Плотная масса рубцового содержимого, таким образом, благодаря моторике рубца медленно вращается против часовой стрелки. Происходящее при этом размельчение и перемешивание кормовой массы способствует созданию оптимальных условий для жизнедеятельности микрофлоры. Прекращение моторики рубца приводит к гибели симбиотической микрофлоры, развитию гнилостных процессов в рубце и сильной интоксикации организма.
Сетка сокращается каждые 30...60 с. Различают две фазы: вначале сетка уменьшается в размерах наполовину, затем слегка расслабляется, после чего сокращается уже полностью. Во время отрыгивания жвачки происходит дополнительное третье сокращение. При сокращении сетки грубые крупные частицы содержимого выталкиваются обратно в рубец, а измельченная и полужидкая пищевая масса поступает в книжку, а затем в сычуг. Переходу пищевой массы из книжки в сычуг способствует то, что во время сокращения сетки сычуг расширяется, в нем создается отрицательное давление и в результате этого жидкая масса засасывается из книжки.
210
14*
211
 Книжка
сокращается в поперечном и продольном
направлениях,
благодаря этому происходит дополнительное
мацериро-вание
задержанных грубых частиц корма. Между
листочками книжки
более грубые частицы корма подвергаются
дальнейшему перевариванию. Нежные,
тонкие разветвленные листочки книжки
обладают
отличной всасывающей способностью. В
книжке интенсивно
всасывается вода, растворенные в ней
органические вещества
и минеральные соли, а уплотненное
содержимое переходит в
сычуг, где снова разбавляется, но уже
желудочным соком, содержащим
соляную кислоту. Таким образом происходит
смена реакции
жидкой фазы содержимого преджелудков.
Книжка
сокращается в поперечном и продольном
направлениях,
благодаря этому происходит дополнительное
мацериро-вание
задержанных грубых частиц корма. Между
листочками книжки
более грубые частицы корма подвергаются
дальнейшему перевариванию. Нежные,
тонкие разветвленные листочки книжки
обладают
отличной всасывающей способностью. В
книжке интенсивно
всасывается вода, растворенные в ней
органические вещества
и минеральные соли, а уплотненное
содержимое переходит в
сычуг, где снова разбавляется, но уже
желудочным соком, содержащим
соляную кислоту. Таким образом происходит
смена реакции
жидкой фазы содержимого преджелудков.
Сокращение преджелудков регулирует находящийся в продолговатом мозге нервный центр. Парасимпатические нервы усиливают, а симпатические — тормозят сокращения преджелудков. На сокращения преджелудков влияет также кора больших полушарий. Раздражение рецепторов ротовой полости при пережевывании корма учащает и усиливает сокращения преджелудков. Раздражение рецепторов двенадцатиперстной кишки тормозит сокращение преджелудков. Отделы преджелудков рефлекторно влияют на моторику друг друга. Например, переполнение сычуга тормозит сокращения книжки, а переполнение книжки тормозит сокращения рубца и сетки. Моторика преджелудков сохраняется и при нарушении их связи с центральной нервной системой за счет интрамуральных нервных сплетений, однако при этом различные отделы сокращаются несогласованно.
Жвачный процесс. Характерной особенностью пищеварительных функций у жвачных животных является наличие процесса жвачки — отрыгивание части плотного содержимого рубца и его повторное пережевывание. Жвачный период начинается спустя некоторое время после еды в зависимости от характера корма и внешних условий: у крупного рогатого скота через 30...70 мин, у овец через 20...45 мин. За это время корм в рубце набухает и частично размягчается, что облегчает его пережевывание. Грубый корм задерживает появление жвачного периода. Быстрее начинается жвачный период при полном покое у лежащего животного. В ночное время жвачные периоды возникают чаще, чем днем. В сутки бывает 6...8 жвачных периодов, каждый из которых длится по 40...50 мин. В течение суток коровы пережевывают до 100 кг содержимого рубца.
В начале отрыгивания возникает дополнительное сокращение сетки и пищеварительного желоба, в результате чего жидкое содержимое сетки поднимается к кардиальному отверстию пищевода. Одновременно происходит остановка дыхания в фазе выдоха, а затем следует попытка вдоха при закрытой гортани. При этом сокращаются мышцы-инспираторы и диафрагма, но воздух в легкие не попадает. В связи с этим давление в грудной полости резко падает до 46...75 мм рт. ст., что приводит к засасыванию разжижен-
ной массы в пищевод. Затем дыхание восстанавливается и антиперистальтические сокращения пищевода способствуют продвижению пищевого кома по пищеводу в ротовую полость. После попадания отрыгиваемой массы в ротовую полость животное мелкими порциями заглатывает жидкую часть, а плотную, оставшуюся в ротовой полости, тщательно пережевывает.
Таким образом, жвачный процесс — это проявление сложного висцеромоторного рефлекса. Возбуждение от тактильных рецепторов преджелудков по центростремительным нервным волокнам в составе вагосимпатических нервов достигает центра жвачки в продолговатом мозге. Отсюда импульсы передаются на центр дыхания и ядра блуждающих нервов, затем по центробежным волокнам в составе блуждающих нервов к мышцам сетки, пищеводного желоба, пищевода, гортани и дыхательным мышцам.
Стимулирует отрыгивание содержимого преджелудков раздражение рецепторов преджелудка, особенно сетки и пищеводного желоба, грубыми частями корма. Пережеванная масса заполняет книжку и переходит в сычуг. Раздражение рецепторов этих отделов тормозит жвачный процесс.
Пищеварение в сычуге. Слизистая оболочка сычуга содержит железы, вырабатывающие сычужный сок. Сычужные железы секрети-руют непрерывно. За сутки образуется довольно большое количество сычужного сока: у коров — 40...80 л, у телок и бычков — 30...40, у взрослых овец —4... 11 л. При каждом кормлении животного происходит усиление секреции. У овец рН сока колеблется от 0,97 до 2,2, у коров — от 1,5 до 2,5. Как и у животных с однокамерным желудком, наиболее важные составляющие сычужного сока — это ферменты (пепсин, химозин, липаза) и соляная кислота.
Одна из существенных особенностей сычужного пищеварения — непрерывная секреция желудочного сока в связи с постоянным поступлением в сычуг предварительно подготовленной однородной массы. Такое состояние сычужных желез поддерживается постоянным раздражением механо- и хеморецепторов самого сычуга и интерорецептивным влиянием преджелудков.
Гуморальная фаза сычужной секреции осуществляется при участии гормонов и метаболитов пищеварительного тракта (гастрин, энтерогастрин, гистамин и др.). В регуляции секреторной деятельности сычуга принимают участие гормоны щитовидной железы, надпочечников, поджелудочной железы, половых желез и др.
В зависимости от вида кормов выделяется разное количество сычужного сока. Наибольшее его количество с высокой кислотностью и переваривающей способностью образуется при скармливании травы и сена бобовых трав, зерновых кормов и жмыха.
Сокращения сычуга, подобно однокамерному желудку, проявляются в форме перистальтических и слабых тонических движений. Периоды сокращений сменяются периодами покоя. Порция содержимого проходит через сычуг за 30...60 мин.
212
213
 Особенности
желудочного пищеварения у молодняка
жвачных.
Особенности
желудочного пищеварения у молодняка
жвачных.
Пищеварительные процессы в желудочно-кишечном тракте новорожденных (телят, ягнят и др.) значительно отличаются от аналогичных процессов у взрослых животных.
У новорожденных жвачных преджелудки недостаточно развиты в морфологическом и функциональном отношении (объем рубца, сетки и книжки соответствует половине сычуга). Интенсивный рост преджелудков происходит в первые месяцы жизни и стимулируется поеданием твердого корма. В годовалом возрасте рубец уже составляет около 80 % объема сложного желудка. У новорожденных телят, питающихся молоком, рубец недостаточно заселен бактериями и не принимает участия в процессе гидролиза и сбраживания питательных субстратов. В связи с этим содержание глюкозы и летучих жирных кислот в крови молодняка жвачных, питающихся молоком, аналогично таковому у животных с однокамерным желудком. По мере развития рубца содержание глюкозы в крови жвачных падает более чем в два раза, но повышается уровень летучих жирных кислот.
У молодняка жвачных молоко, минуя рубец, переходит из пищевода в книжку по так называемому пищеводному желобу, образованному складками слизистой оболочки. Главным стимулом для рефлекторного смыкания пищеводного желоба служит сам акт сосания. Рецепторы, раздражение которых вызывает этот рефлекс, находятся в слизистой оболочке языка, губ и глотки (рис. 5.8). При медленной выпойке желоб смыкается полностью и проглоченное молоко поступает непосредственно в сычуг. При быстром заглатывании большого количества пищеводный желоб смыкается не полностью и молоко частично попадает в рубец, где может задерживаться и загнивать. Поэтому телятам-молочникам молоко необходимо выпаивать из поилок с соской. Рефлекс пищеводного желоба сохраняется у телят до 2-месячного возраста, а затем постепенно угасает. По мере развития рубца в нем формируется популяция микроорганизмов, попадающих с кормом и водой. Важными факторами, участвующими в этом процессе, являются контакт телят со взрослыми животными и начало приема растительного корма. В рубце 2...3-месячных животных целлюлозо-литическая активность уже достаточно высокая.
//

Рис. 5.8. Схема рефлекторной регуляции смыкания пищеводного желоба:
1 — рецепторы ротовой полости; 2— афферентный путь от рецепторов ротовой полости; J — нервный центр продолговатого мозга; 4— эфферентный путь в составе блуждающих нервов; 5—пищеводный желоб; б—сетка; 7—рубец; 8— книжка; 9— сычуг; 10— пищевод; 11 — высшие центры рефлекса пищеводного желоба
В первые сутки у новорожденных отсутствуют жвачные периоды. У телят и ягнят они впервые появляются в возрасте 9... 10 сут. Вначале жвачные периоды возникают редко (2...4 раза в сутки) и длятся всего 2...8 мин, затем их частота и продолжительность возрастают.
Сычуг также претерпевает определенные изменения в ходе онтогенеза. К моменту рождения все слои стенки сычуга оформлены, однако его железы не достигают полного развития. У новорожденных телят большая часть соляной кислоты находится в сычуге в связанном состоянии. В первые дни после рождения в желудочном соке мало ферментов, поэтому важную роль в гидролизе питательных веществ играют ферменты материнского молока. С возрастом в сычужном соке увеличивается кислотность и концентрация активных протеолитических ферментов, причем у телят, получающих молочную пищу, преобладают химозин и желудочная липаза.