

3.1.2. Механизм сокращения мышцы
Механизм мышечного сокращения на молекулярном уровне объясняет теория скользящих нитей, разработанная Хаксли и Хансон (1954).
При электронной микроскопии обнаружено, что на миозиновых нитях имеются выступы, получившие название поперечных мостиков. Поперечные мостики, состоящие из головки и шейки, протянувшиеся от миозиновых нитей, в спокойном состоянии не могут соединиться с актиновыми нитями из-за особого расположения тропомиозина, закрывающего активные центры актина и препятствующего их взаимодействию с поперечными мостиками миозина. Тропонин подавляет миозин-АТФазную активность, что делает невозможным расщепление АТФ, в результате мышечные волокна пребывают в расслабленном состоянии (рис.11).
С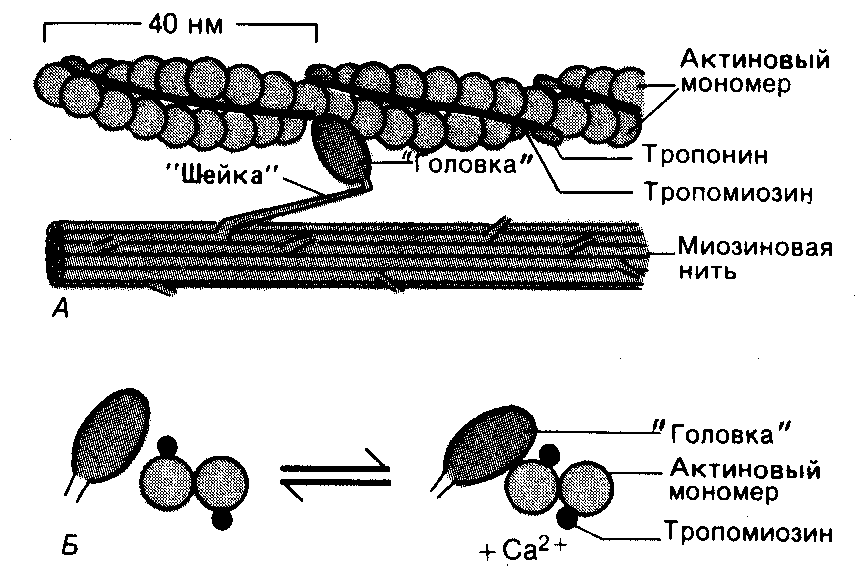 окращение
скелетной мышцы волокон начинается с
приходом нервного импульса к сарколемме,
который приводит к формированию ПД.
Возникающий ПД распространяется по
поверхностной мембране, а также по
мембранам, выстилающим поперечные
трубочки Т-системы. Проникая внутрь
волокна, электрическая волна приводит
к деполяризации мембран продольных
трубочек и цистерн саркоплазматического
ретикулума. Деполяризация мембраны
цистерн открывает электровозбудимые
кальциевые каналы. В связи с тем, что в
саркоплазме концентрация кальция менее
10-7 М/л, а в саркоплазматическом
ретикулуме − более 10-4 М/л,
начинается интенсивный выход ионов
кальция в саркоплазму. Выделившийся
свободный Са2+ и является
инициатором мышечного сокращения.
окращение
скелетной мышцы волокон начинается с
приходом нервного импульса к сарколемме,
который приводит к формированию ПД.
Возникающий ПД распространяется по
поверхностной мембране, а также по
мембранам, выстилающим поперечные
трубочки Т-системы. Проникая внутрь
волокна, электрическая волна приводит
к деполяризации мембран продольных
трубочек и цистерн саркоплазматического
ретикулума. Деполяризация мембраны
цистерн открывает электровозбудимые
кальциевые каналы. В связи с тем, что в
саркоплазме концентрация кальция менее
10-7 М/л, а в саркоплазматическом
ретикулуме − более 10-4 М/л,
начинается интенсивный выход ионов
кальция в саркоплазму. Выделившийся
свободный Са2+ и является
инициатором мышечного сокращения.
Рис.11. Действие Са2+ во время активации миофибриллы.
А. Актиновая и миозиновая нити на продольном сечении волокна.
Б. Они же на поперечном сечении. Когда Са2+ связывается с тропонином, тропомиозин попадает в желобок между двумя мономерами актина, обнажая участки прикрепления поперечных мостиков (по Haxley, 1973 c изменениями Р.Шмидта и Г. Тевса, 1995).
Достаточный для начала мышечного сокращения уровень ионов кальция достигается через 12-15 мс после прихода нервного импульса. Это скрытый, латентный период мышечного сокращения. В связи с тем, что скорость распространения ПД по сарколемме выше времени, необходимого для выделения Са2+из саркоплазматического ретикулума, то все фибриллы участка мышцы, иннервируемого одним нервом, сокращаются одновременно.
С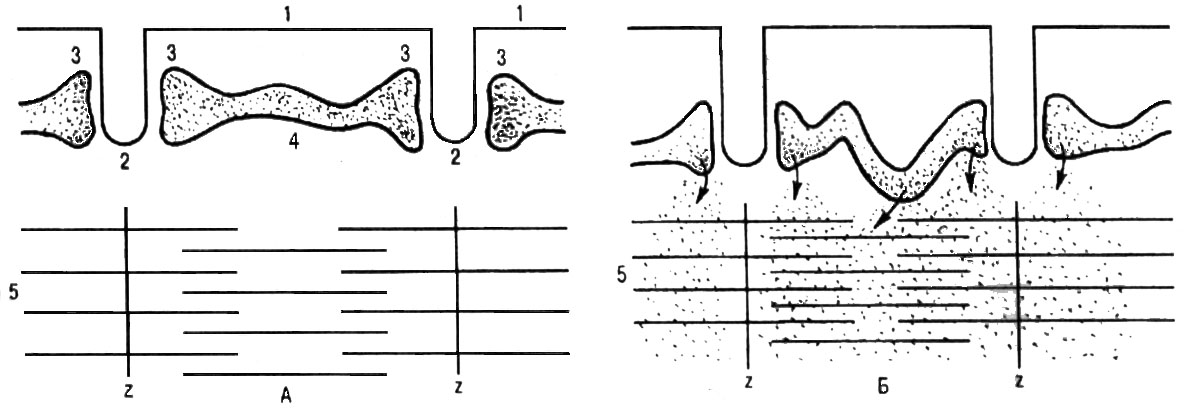 овокупность
явлений, обусловливающих связь между
возбуждением (потенциалом действия) и
сокращением мышечных волокон, получила
название «электромеханического
сопряжения», или «электромеханической
связи».
овокупность
явлений, обусловливающих связь между
возбуждением (потенциалом действия) и
сокращением мышечных волокон, получила
название «электромеханического
сопряжения», или «электромеханической
связи».
Рис. 12. Взаимоотношения клеточной мембраны (1), поперечных трубочек (2), боковых цистерн (3) и продольных трубочек (4) саркоплазматического ретикулума, миозиновых и актиновых нитей (5) мышечного волокна. А—в состоянии покоя; Б—во время сокращения. Деполяризация мембраны и поперечных трубочек вызвала освобождение ионов Са2+ из боковых цистерн. Освободившиеся ионы Са2+ диффундируют по направлению к миофибриллам и частично захватываются продольными трубочками ретикулума.
Механизм инициации сократительного процесса представляется в настоящее время следующим образом. В присутствии ионов Са2+, а также АТФ тропонин изменяет свою конфигурацию и отодвигает нить тропомиозина, открывая возможность соединения головки поперечного мостика миозина с актином (см. рис.11). Соединение головки фосфорилированного миозина с актином приводит к тому, что головка приобретает АТФазную активность, в ней происходит гидролиз АТФ, сопровождаемый изменением пространственной ориентации. Последнее носит форму гребкового движения, обеспечивающего втягивание тонких актиновых миофиламентов в промежутки между толстыми миозиновыми миофиламентами на один шаг (примерно 20 нм) без изменения длины миофиламентов. После этого, образующиеся Ф и АДФ, отходят, а на их место присоединяется новая молекула АТФ. Это приводит к разрыву связи миозина с активным центром актина. Затем следуют отрыв мостика и повторение всего цикла. При каждом гребковом движении головки поперечного мостика расщепляется одна молекула АТФ. Скорость расщепления АТФ является фактором, предопределяющим частоту гребковых движений и, таким образом, скорость скольжения нитей актина относительно нитей миозина. Целая мышца укорачивается в результате сокращения множества саркомеров, соединенных последовательно в миофибриллах (рис. 12).
Обнаружено, что при сокращении скелетной мышцы лягушки поперечные мостики должны совершить за 0,1 с 50 гребковых движений, чтобы обеспечить укорочение каждого саркомера волокна на 50 %.
Таким образом, при сокращении мышцы:
− актиновые и миозиновые нити практически не укорачиваются;
− взаимодействие актина с миозином приводит к взаимному вхождению нитей в промежутки между ними;
− в результате две соседние Z-мембраны приближаются друг к другу и при максимально сильном сокращении расстояние между ними может уменьшиться почти в два раза;
− так как заключенная внутри саркомера саркоплазма несжимаема, то при уменьшении длины саркомер (мышца) расширяется;
− подобные процессы одновременно протекают во всех саркомерах мышечного волокна, поэтому оба конца мышцы подтягиваются к центру.
Расслабление мышцы. Гребковые движения («шаги») поперечных мостиков будут повторяться до тех пор, пока в саркоплазме есть свободный Са2+ (в концентрации более 10-5) и АТФ. Если нет новой волны деполяризации, то кальций быстро убирается обратно в цистерны саркоплазматического ретикулума. Работа насоса активируется самим кальцием, вернее возрастанием его концентрации в саркоплазме. Работа насоса требует затраты большого количества АТФ: для удаления каждого иона кальция используется 2 молекулы АТФ. Результатом откачивания кальция из саркоплазмы является разрыв всех связей актина и миозина и расслабление мышцы.
В целом последовательность событий в цикле сокращение—расслабление мышечного волокна представляется в следующем виде:
— поступление ПД по нервному волокну к мионевральному синапсу;
— синаптическая активация мышечного волокна;
— возникновение ПД, проведение его вдоль клеточной мембраны и в глубь волокна по Т-трубочкам;
— освобождение ионов Са2+ из боковых цистерн саркоплазматического ретикулума, диффузия его к миофибриллам;
— конформация тропонин-тропомиозинового комплекса;
— контакт поперечных мостиков миозина с актином;
— освобождение энергии АТФ;
— скольжение актиновых и миозиновых нитей, выражающееся в укорочении миофибриллы;
— активация кальциевого насоса;
— снижение концентрации свободных ионов Са2+ в саркоплазме;
— расслабление миофибрилл.