

3.1.1. Структурно-функциональная характеристика
Структурной и сократительной единицей мышцы является мышечное волокно, представляющее собой сильно вытянутую многоядерную клетку. Толщина волокна — в пределах 10—100 мкм, чаще всего около 50 мкм. Длина волокна определяется размером мышцы и может составлять от нескольких миллиметров до нескольких сантиметров.
У человека количество мышечных волокон устанавливается через 4—5 месяцев после рождения и затем практически не меняется. При рождении ребенка их диаметр составляет примерно 1/5 толщины волокон взрослых людей. Диаметр мышечных волокон может значительно увеличиваться под воздействием тренировки (функциональная гипертрофия).
Основными структурно-функциональными элементами мышечной клетки являются:
− мембранная оболочка;
− саркоплазма с органоидами;
− миофиламенты.
Как и всякая клетка, мышечное волокно имеет мембранную оболочку — сарколемму. Мембрана мышечного волокна образована типичной плазматической мембраной, укрепленной соединительно-тканными волокнами. Эти волокна объединяясь у концов мышечных волокон образуют сухожилия, с помощью которых мышца крепится к костям.
Р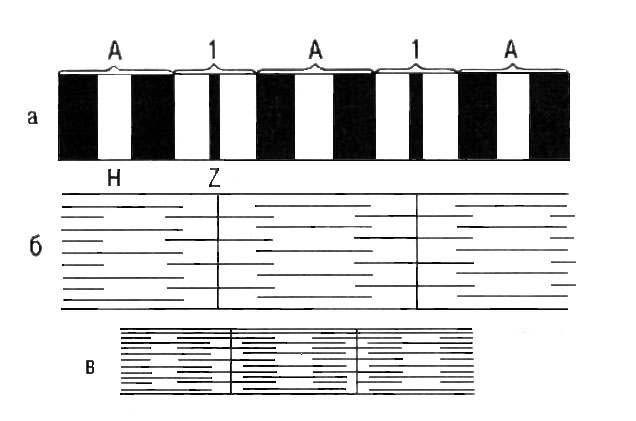 ис.
10. Структура миофибрилл.
Показаны диски А и I, полоски Z
и Н (а). Взаимное расположение толстых
(миозиновых) и тонких (актиновых) нитей
в расслабленной (б) и сокращенной (в)
миофибрилле.
ис.
10. Структура миофибрилл.
Показаны диски А и I, полоски Z
и Н (а). Взаимное расположение толстых
(миозиновых) и тонких (актиновых) нитей
в расслабленной (б) и сокращенной (в)
миофибрилле.
В саркоплазме (цитоплазме) мышечного волокна находится типичный набор органоидов: многочисленные ядра, митохондрии, саркоплазматический ретикулум (см. ниже), растворимые белки (миоглобин), капельки жира, гранулы гликогена, фосфатсодержащие вещества, другие малые молекулы, ионы.
От одного конца мышечного волокна до другого тянется масса нитей — миофибрилл, с которыми связана способность мышцы к сокращению. Их диаметр составляет 1— 2 мкм, так что в одном мышечном волокне может насчитываться более 2000 миофибрилл. Миофибриллы группируются в пучки, по 4—20 штук в каждом. В одиночной миофибрилле насчитывается 2000—2500 протофибрилл — толстых и тонких.
Они располагаются параллельно друг другу внутри мышечной клетки. Миофибриллы представляют собой сократимые пучки “нитей” (филаментов) диаметром около 1 мкм. Перегородки, называемые Z-пластинками, разделяют их на несколько компартментов – саркомеров − длиной примерно по 2,5 мкм. Структура саркомеров схематически показана на рис. С помощью светового микроскопа в них можно видеть регулярно чередующиеся поперечные светлые и темные полосы. Согласно теории Хаксли и Хансон [1954], такая поперечная полосатость миофибрилл обусловлена особым взаиморасположением актиновых и миозиновых филаментов. Середину каждого саркомера занимают несколько тысяч толстых нитей миозина диаметром примерно по 10 нм. На обоих концах саркомера находятся около 2000 тонких (толщиной по 5 нм) нитей актина, прикрепленных к Z-пластинкам наподобие щетинок в щетке. Пучок лежащих в определенном порядке миозиновых нитей длиной 1,6 мкм в середине саркомера выглядит в световом микроскопе темной полосой шириной 1,6 мкм. Из-за свойства двойного лучепреломления в поляризованном свете (т. е. анизотропии) она называется А-диском. По обе стороны от А-диска находятся изотропные участки, содержащие только тонкие нити и поэтому выглядящие светлыми; эти так называемые I-диски тянутся до Z-пластинок.
Светлые I-диски делятся пополам поперечной темной Z-линией, с обеих сторон которой крепятся тонкие (актиновые) протофибриллы. Участок между двумя соседними Z-линиями называется саркомером. Он представляет собой основную повторяющуюся структуру миофибриллы. Длина саркомера обычно лежит в пределах 2—3 мкм. Эта зона перекрывания в А-диске выглядит в световом микроскопе гораздо темнее центральной Н-зоны, в которой нет актиновых нитей. На электронных микрофотографиях Н-зоны видна очень тонкая темная М-линия в середине саркомера − сеть опорных белков, по-видимому, удерживающих толстые нити в составе единого пучка.
При длине типичного мышечного волокна человека 5 см его миофибриллы насчитывают порядка 20 000 последовательно расположенных саркомеров. Миофибриллы, точнее их саркомеры, являются сократительным аппаратом, «мотором» мышечного волокна.
Итак, в каждом саркомере имеется два набора тонких нитей, прикрепленных к Z-ли-ниям, и один комплект толстых нитей, сосредоточенных в А-диске. В покоящейся мышце концы толстых и тонких филаментов лишь незначительно перекрываются на границе между А- и I-дисками. При сокращении мышцы зона перекрытия тонких и толстых филаментов увеличивается, ширина А-дисков и Н-зон уменьшается (см. рис. 10 ).
Один грамм ткани скелетной мышцы содержит около 100 мг сократительных белков. В настоящее время в основных чертах установлена тонкая структура тонких и толстых филаментов. Толстые филаменты состоят из сократительного белка миозина, а тонкие — из актина.
Миозиновые филаменты образуются более чем 200 молекулами миозина (молекулярная масса 42000). Каждая из них скручена попарно и имеет выступающий отросток, называемый головкой. Головки направлены под углом от центра в сторону тонких нитей (напоминают ёрш для мытья посуды). В основании головки миозина имеется фермент АТФ-аза, а на самой головке располагаются легкие цепи и молекула АТФ.
Актиновые филаменты скомпанованы из двух актиновых нитей, представляющих собой как бы бусинки глобулярных молекул актина (молекулярная масса 500000). Тонкие нити имеют активные центры, расположенные друг от друга на расстоянии 40 нм, к которым могут прикрепляться головки миозина. Кроме актина в тонких нитях имеются и другие белки − тропомиозин, тропонины (I,T,C). Тропомиозин и тропонин образуют регуляторный белковый комплекс, который в невозбужденной мышце блокирует взаимодействие между актином и миозином. В результате невозбужденная мышца расслаблена. Блокирующая роль тропонина снимается ионами кальция (Са2+), поступающими к сократительным белкам при возбуждении.
В поперечном сечении толстые и тонкие филаменты расположены правильными шестиугольниками так, что каждая толстая нить окружена шестью тонкими, а каждая тонкая может вступать в контакт с тремя толстыми.
Система микротрубочек скелетной мышцы представляет собой сложный биокомплекс. Во многих участках поверхностная мембрана мышечной клетки образует углубления в виде Т-трубочек (диаметром 50 нм), перпендикулярных продольной оси волокна; эта система поперечных трубочек соединяется с внеклеточной средой и обычно окружает каждую миофибриллу в области I-дисков (у высших позвоночных ).
Перпендикулярно поперечным трубочкам, т.е. параллельно миофибриллам, расположена система продольных трубочек (истинный саркоплазматический ретикулум). Пузырьки на их концах (терминальные цистерны) прилегают к мембранам ситемы поперечных трубочек, образуя так называемые триады. В этих пузырьках и хранится внутриклеточный кальций. В отличие от поперечной системы продольная не сообщается с внеклеточной средой. Саркоплазматический ретикулум выполняет важную функцию в инициации сокращения мышцы, являясь депо Са2+. Мембраны саркоплазматического ретикулума содержат работающий на энергии АТФ кальциевый насос, который осуществляет активный транспорт Са2+ из миоплазмы в продольные трубочки, снижая таким образом примерно до 10-7 М миоплазматическую концентрацию этих ионов в покоящейся (расслабленной) мышце.