

4. Биологическое разнообразие. Генетический полиморфизм популяций как основа биологического разнообразия. Проблема сохранения биоразнообразия
Под биологическим разнообразием понимается все «множество различных живых организмов, изменчивость среди них и экологических комплексов, частью которых они являются, что включает разнообразие внутри видов, между видами и экосистемами»; при этом следует различать глобальное и локальное разнообразие. Биологическое разнообразие представляет собой один из важнейших биологических ресурсов (биологическим ресурсом считается «генетический материал, организмы или их части, либо экосистемы, используемые или потенциально полезные для человечества, включая природное равновесие внутри экосистем и между ними»).
Различают следующие типы биологического разнообразия: альфа, бета, гамма и генетическое разнообразие. Под α -разнообразием понимают видовое разнообразие, под β-разнообразием – разнообразие сообществ на определенной территории; γ-разнообразие – это интегральный показатель, включающий α- и β-разнообразие. Однако в основе перечисленных типов биоразнообразия лежит генетическое (внутривидовое, внутрипопуляционное) разнообразие.
Наличие двух и более аллелей (а, соответственно, и генотипов) в популяции называетсягенетическим полиморфизмом. Условно принято, что частота наиболее редкого аллеля при полиморфизме должна быть не менее 1% (0,01). Существование генетического полиморфизма – обязательное условие сохранения биоразнообразия.
Представления о необходимости сохранения генетического полиморфизма в природных популяциях были сформулированы еще в 1920-ых гг. нашими выдающимися соотечественниками. Николай Иванович Вавилов создал учение об исходном материале, обосновал необходимость создания хранилищ мирового генофонда культурных растений. Александр Сергеевич Серебровский создал само учение о генофонде. В понятие «генофонд» вкладывалось то генетическое разнообразие вида, которое сложилось в ходе его эволюции или селекции и обеспечило его адаптивные и продукционные возможности. Сергей Сергеевич Четвериков заложил основы учения и методов оценки генетической гетерогенности популяций диких видов растений и животных.
Глобальные экологические проблемы обострились после Второй мировой войны. Для их решения в 1948 г. был образован Международный союз охраны природы и природных ресурсов(МСОП). Первоочередной задачей МСОП являлось составление Красных книг – списков редких и находящихся под угрозой исчезновения видов. В 1963-1966 гг. была издана перваяМеждународная Красная книга. В 1980 г. вышло ее четвертое издание. В 1978-1984 гг. издается Красная книга СССР, а в 1985 г. – Красная книга Российской Федерации.
Однако человечество осознало всю серьезность этой проблемы лишь в последней четверти XX века. Чуть более тридцати лет назад (1972 г.) в Стокгольме состоялась первая конференция ООН по окружающей человека природной среде. На этом форуме были намечены общие принципы международного сотрудничества в области охраны природы. На основе решений Стокгольмской конференции были сформулированы современные принципы сохранения среды жизни.
Первый принцип – это принцип всеобщей связи в живой природе: выпадение одного звена в сложной цепи трофических и иных связей в природе может привести к непредвиденным результатам. Этот принцип основан на классических представлениях о существовании причинно-следственных связей между элементами надорганизменныхбиологических систем, причем, множество этих связей приводит к образованию различных цепей, сетей и пирамид.
Отсюда вытекает принцип потенциальной полезности каждого компонента живой природы: невозможно предвидеть, какое значение для человечества будет иметь тот или иной вид в будущем. В общественном сознании утрачивает значение разграничение видов на «полезные» и «вредные», утверждается представление о том, что «вредный или сорный вид – это всего лишь организм не на своем месте».
На основании принципов всеобщей связи и потенциальной полезности каждого компонента живой природы формируется концепция невмешательства в процессы, протекающие в природных экосистемах: «Мы не знаем, к чему это приведет, поэтому лучше оставить все как есть». Идеальным способом сохранить status quo считалось создание охраняемых территорий с абсолютным заповедным режимом. Однако практика заповедного дела показала, что современные экосистемы уже утратили способность к естественному самовосстановлению, и для их сохранения требуется активное вмешательство человека.
В результате стал неизбежным переход от концепции невмешательства и консервации существующего положения к концепции устойчивого развития общества и биосферы. Концепция устойчивого развития подразумевает повышение экологического и ресурсного потенциала природных экосистем, создание устойчивых контролируемых экосистем, удовлетворение потребностей общества в природных ресурсах на основе научно обоснованного рационального, неистощительного и многоцелевого природопользования, охраны, защиты и воспроизводства всех компонентов экосистем.
Дальнейшее развитие концепции устойчивого развития неизбежно привело к принципу необходимости сохранения биологического разнообразия: только многообразная и разнообразная живая природа оказывается устойчивой и высокопродуктивной. Принцип необходимости сохранения биологического разнообразия в полной мере согласуется с основными принципами биоэтики: «каждая форма жизни уникальна и неповторима», «каждая форма жизни имеет право на существование», «что не нами создано, не должно быть нами разрушено». При этом ценность генотипа определяется не его полезностью для человека, а его уникальностью. Таким образом, было признано, что «сохранение генофонда есть ответственность перед дальнейшей эволюцией» (Франкель, XIII Международный генетический прогресс в Беркли, 1974). Сваминатан (Индия) выделил три уровня ответственности за сохранение генофонда: профессиональный, политический и общественный.
В 1980 г. Международным союзом охраны природы и природных ресурсов была разработана «Всемирная стратегия охраны природы». В материалах Всемирной стратегии отмечается, что одной из глобальных экологических проблем является проблема питания: 500 млн. человек систематически недоедают. Труднее учесть число людей, не получающих полноценного питания, сбалансированного по белкам, витаминам и микроэлементам.
Всемирная стратегия сформулировала первоочередные задачи охраны природы:
– поддержание главных экологических процессов в экосистемах.
– Сохранение генетического разнообразия.
– Долгосрочное рациональное использование видов и экосистем.
В 1992 г. в Рио-де-Жанейро на конференции Организации Объединенных Наций по окружающей среде и развитию (UNCED) был принят ряд документов, подписанных представителями 179 государств:
– Программа действий: Повестка дня на XXI век.
– Заявление о принципах в отношении лесов.
– Конвенция ООН об изменении климата.
– Конвенция о биологическом разнообразии.
В материалах Конвенции о биологическом разнообразии отмечается, что «...разнообразие важно для эволюции и сохранения систем жизнеобеспечения биосферы». Для сохранения систем жизнеобеспечения биосферы необходимо сохранение всех форм биологического разнообразия: «Страны, которые присоединяются к Конвенции, должны определять компоненты биологического разнообразия, ...осуществлять контроль за видами деятельности, которые могут оказать вредное воздействие на биологическое разнообразие».
На конференции UNCED было признано, что снижение уровня биологического разнообразия является одной из основных причин прогрессирующей деградации природных экосистем. Не вызывает сомнения, что только при условии сохранения оптимального уровня разнообразия возможно создание экосистем, устойчивых к экстремальным воздействиям физико-химических факторов, вредителей и болезней.
В 1995 г. в Софии на конференции министров окружающей среды европейских стран была принята Общеевропейская стратегия сохранения биологического и ландшафтного разнообразия. Перечислим принципы Общеевропейской стратегии сохранения биологического и ландшафтного разнообразия природы:
– Охрана наиболее уязвимых экосистем.
– Охрана и восстановление нарушенных экосистем.
– Охрана территорий с наибольшим видовым разнообразием.
– Сохранение эталонных природных комплексов.
Прекращение роста продуктивности искусственных экосистем также связывается с низким уровнем биоразнообразия: в настоящее время возделывается всего 150 видов культурных растений и разводится 20 видов домашних животных. При этом низкий уровень глобального разнообразия сочетается и с низким уровнем локального разнообразия, с господством монокультуры или культурооборотов с коротким сроком ротации. Погоня за однородностью сортов растений и пород животных привела к резкому сужению генетического разнообразия. Следствием снижения разнообразия является снижение устойчивости к экстремальным физико-химическим факторам среды и, в еще большей степени, к вредителям и болезням.
Многочисленными исследованиями доказано, что единственный надежный путь к повышению устойчивости и продуктивности природных экосистем – это повышение уровня их гетерогенности, поскольку в генетически неоднородных системах возникают компенсаторные взаимодействия особей с различными особенностями роста и развития, чувствительности к динамике факторов среды, болезням, вредителям. Именно гетерогенные насаждения обеспечивают возможность непрерывного и неистощительного природопользования.
Следовательно, назрела необходимость более широкого использования видового и внутривидового (генетического) потенциала возможно большего числа видов, пригодных для культивирования в контролируемых условиях. Все разнообразие материала, подлежащего сохранению, включает следующие категории организмов: сорта и породы, возделываемые и разводимые в настоящее время; сорта и породы, вышедшие из производства, но представляющие большую генетическую и селекционную ценность по отдельным параметрам; местные сорта и аборигенные породы; дикие сородичи культурных растений и домашних животных; дикие виды растений животных, перспективные для введения в культуру и доместикации; экспериментально созданные генетические линии.
Естественно, что для решения комплекса задач, связанных с проблемами биологического разнообразия, требуется вначале выработать критерии для оценки биоразнообразия, выявить и оценить уровень разнообразия в конкретных экосистемах (природно-территориальных комплексах), разработать рекомендации по сохранению и приумножению выявленного разнообразия, апробировать и внедрить эти рекомендации в агропромышленное производство.
12. Популяционные волны как эволюционный фактор.
Популяционные волны как элементарный эволюционный фактор
При характеристике популяции как элементарной эволюционной единицы одной из важных и постоянных черт признано явление колебания ее численности — волн жизни (популяционные волны, см. рис. 7.1). Популяционные волны имеют важное значение как один из элементарных факторов эволюции.
Периодические или апериодические колебания численности характерны для всех без исключения видов живых организмов. Конкретные причины таких флуктуации могут быть бесконечно разнообразны (как биотические, так и абиотические).
Классификация популяционных волн. В природе встречается множество популяционных волн. Рассмотрим основные.
Периодические колебания численности короткоживущих организмов характерны для большинства насекомых, однолетних растений, большинства грибов и микроорганизмов. В наиболее простом виде это сезонные колебания численности.
2. Непериодические колебания численности, зависящие от сложного сочетания разных факторов. В первую очередь они зависят от благоприятных для данного вида (популяции) отношений в пищевых цепях: ослабление пресса хищников для популяций жертв или, например, увеличение кормовых ресурсов для популяций хищников.
Обычно такие колебания численности касаются не одного-двух, а многих видов животных и растений в биогеоценозах и порой ведут к коренным перестройкам всего биогеоценоза.
3. Вспышки численности видов в новых районах, где отсутствуют их естественные враги. Примерами колебаний численности в XIX—XX вв. служат кролики в Австралии, домовые воробьи в Северной Америке, канадская злодея, американская норка и ондатра в Евразии и др. В XVI—XVII вв. вспышки численности испытывали расселявшиеся с мореплавателями по всему миру крысы (Rattus norvegicus). С возникновением городов неимоверно увеличилась численность домовой мухи (Musca domestica) в связи с распространением помоек и гниющих пищевых запасов около поселений человека.
4. Резкие непериодические колебания численности, связанные с природными «катастрофами» (разрушением биогеоценозов или целых ландшафтов). Несколько засушливых лет могут быть причиной серьезных изменений в облике больших территорий (наступление луговой растительности на болотистые места, увеличение площади сухих лугов, выгорание большогочисла тор-фяниковых залежей, сохранявшихся на протяжении нескольких тысяч лет как устойчивый биотоп, и т. д.). Виды с подвижными особями (крупные млекопитающие, насекомые, птицы) или живущими в глубоких слоях почвы страдают меньше неподвижных и малоподвижных форм, живущих в лесу и почве. Другие формы гибнут в огромном числе (растения, моллюски, рептилии, амфибии и др.). На пожарищах хорошо известны вспышки численности иван-чая (Epilobium angustifolium) и связанного с ним сообщества насекомых.
Масштабы колебаний численности могут быть различными. Близкие, по-видимому, к максимальным величины колебаний численности в 1 млн раз отмечены в Зауралье для майских жуков (Ю.И. Новожено-вым) и в 12 млн раз — для сибирского шелкопряда в западносибирских лесных биоценозах (А.С. Исаевым).
Эволюционное значение популяционных волн. Если численность какой-либо популяции резко идет на убыль, то от многочисленной популяции случайно могут остаться немногие особи. Например, лесной пожар не затронул небольшой участок леса, где и сохранились остатки популяции короедов (биологические свойства оставшихся в живых короедов в данном случае не имели значения, их спасла случайность) В популяции, пережившей катастрофическое сокращение численности, частоты генов (точнее, частоты аллелей) скорее всего будут иными, чем во всей исходной популяции до начала спада численности. Если вслед за сокращением численности следует резкий ее подъем, то новой вспышке численности дает начало оставшаяся немногочисленная группа. Генотипический состав этой группы и определит новую генетическую структуру всей популяции в период следующего расцвета ее численности. При этом некоторые ранее присутствовавшие в малых концентрациях мутации могут совершенно исчезнуть из популяции, а концентрация других мутаций может случайно резко повыситься (рис. 9.3). В целом произойдет случайное колебание концентраций разных генотипов и мутаций в популяции. В этом и состоит основное эволюционное значение волн численности. Действие волн жизни как эволюционного фактора предполагает неизбирательное, случайное уничтожение особей, благодаря чему редкий перед колебанием численности генотип (аллель) может сделаться обычным и подхвачен в дальнейшем естественным отбором.
При определенных условияхэто случайное и кратковременное колебание численности может превратиться в элементарное эволюционное явление — изменение на протяжении ряда поколений генотипического состава популяции.
Механизм действия популяционных волн в эволюции иллюстрируется модельным примером. В закрытый ящик положено 500 белых и 500 черных шаров, что соответствует частоте аллелей в популяции: Р —0,50. Удалим из ящика наугад 10 шаров (что соответствует «выжившим» 5 диплоидным особям), предположим 4 из них будут черными, 6 — белыми (Р —0,40 и Р —0,60). В следующий момент жизни нашей «популяции» общую численность «особей» увеличим в 100 раз (400 черных, 600 белых). Если мы опять наугад удалим 10 шаров, то велика вероятность, что у нас в руках окажутся шары в соотношении, очень далеком от первоначального (например, 2 черных и 8 белых и соответственно Р —0,20 и Р —0,80). Это соотношение частот в третьей наугад взятой выборке из средней величины «популяции» (200 черных и 800 белых) легко может случайно сдвинуться в сторону теперь уже преобладающих белых, и «популяция» будет представлена почти лишь белыми шарами: велика вероятность того, что мы вытащим 9 белых и 1 черный (Р —0,10 и Р —0,90) Точно так же и в природных популяциях случайные колебания частот аллелей могут быть причиной устранения или резкого повышения концентрации какого-либо гена.
Теоретические расчеты показывают, что влияние популяционных волн может быть особенно заметно в популяциях очень малой величины обычно при численности размножающихся особей не более 500. Именно в этих условиях популяционные волны могут как бы подставлять под действие естественного отбора редкие мутации (внося их в увеличенных концентрациях в популяционный генофонд) или устранять уже довольно обычные варианты.
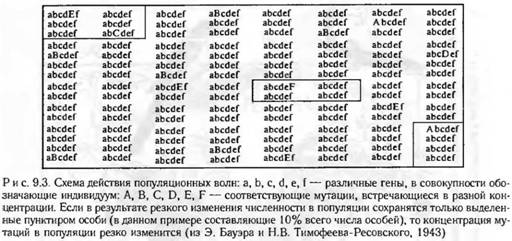
Итак, действие популяционных волн, так же как и действие мутационного процесса, статистично и ненаправленно (хотя природа действия этих эволюционных факторов совершенно различна). Популяционные волны служат «поставщиком» эволюционного материала. Действие этого фактора трудно вычленить в «чистом виде» — очень мало точных данных по оценке величины его давления в природных популяциях. Вероятно, давление популяционных волн, особенно в малых по численности популяциях, должно заметно превышать давление мутационного процесса.
Популяционные волны и мутационный процесс даже при совместном действии еще не могут обеспечить протекания эволюционного процесса. Для этого нужны факторы, длительно действующие в одном направлении. Один из них — изоляция.
13. Изоляция как эволюционный фактор.
Изоляция — возникновение любых барьеров, ограничивающих панмиксию. Значение изоляции в процессе эволюции и сводится к нарушению свободного скрещивания, что ведет к увеличению и закреплению различий между популяциями и отдельными частями всего населения вида. Без такого закрепления эволюционных различий невозможно никакое формообразование.
Разнообразие форм и проявлений изоляции в природе так велико, что для понимания эволюционной роли изоляции необходимо кратко описать основные ее проявления в природе.
Классификация явлений изоляции. В природе существуют пространственная и биологическая изоляции.
Пространственная изоляция может существовать в разных формах: водные барьеры разделяют население «сухопутных» видов, а барьеры суши изолируют население видов-гидробионтов; возвышенности изолируют равнинные популяции, а равнины — горные популяции и т. д. Сравнительно малоподвижные животные — наземные моллюски на Гавайских островах связаны с влажными долинами. В результате, в каждой из сотен долин на больших островах возникает самостоятельная популяция со своими специфическими особенностями. Возникновение территориально-механической изоляции объясняется историей развития видов на определенных территориях. В приведенных двух случаях (рис. 9.4) основной причиной изоляции стало наступление ледников. За время, прошедшее после исчезновения ледников, изолированные формы еще не приобрели значительных морфофизиологических различий и относятся, по-видимому, к единым видам. Впрочем молекулярно-генетические исследования могут показать, что это формы уже возникшего видового ранга (полной изоляции на протяжении жизни десятков тысяч поколений обычно бывает достаточно для возникновения видов).
В настоящее время в связи с деятельностью человека в биосфере все чаще и чаще возникает подобная пространственная изоляция отдельных популяций внутри очень многих видов. Типичным примером стало возникновение в Евразии к началу XX в. разорванного ареала у соболя (Maries zibellina) — результат интенсивного промысла (рис. 9.5). Обычно быстрое возникновение подобного разорванного ареала служит опасным симптомом возможного исчезновения вида.
Пространственная изоляция может возникнуть внутри видов малоподвижных животных и растений, не разделенных заметными физико-географическими барьерами Известно, что обыкновенный соловей (Luscinia luscinia), населяющий многие районы центральной части европейской территории России, в настоящее время практически сплошь находит подходящие условия для гнездования как в не обжитых человеком местах, так и в зарослях по обочинам дорог, в парках и даже скверах больших городов.
При этом наблюдается четко выраженная клинальная изменчивость пения этих птиц: по числу «коленец», по тембру и другим особенностям существуют постепенные переходы от одной местности к другой (характер песни наследственно обусловлен). Возникновение такой клинальной изменчивости возможно только потому, что соловьи, несмотря на существование у них частых сезонных перелетов, обладают большим гнездовым консерватизмом: молодые возвращаются практически на то же место, где они вывелись.
Пространственная изоляция внутри вида существует в двух проявлениях: изоляция какими-либо барьерами между частями видового населения и изоляция, определяемая большей возможностью спаривания близко живущих особей, т. е. изоляция расстоянием. Возникновение пространственной изоляции связано с радиусом репродуктивной активности вида (см. гл. 7).
Радиус репродуктивной активности наземных моллюсков обычно равен нескольким десяткам метров, а чирков — тысяче километров. Значение физико-географических барьеров в пространственной изоляции связано с биологическими особенностями вида в целом.
Биологическую изоляцию обеспечивают две группы механизмов: устраняющие скрещивание (докопуляционные) и изоляция при скрещивании (послекопуляционные). Первые механизмы предотвращают потерю гамет, вторые связаны с потерей гамет и зигот (Э. Майр).
Спариванию близких форм препятствуют различия во время половой активности и созревания половых продуктов. Известно существование «яровых» и «озимых» рас у миног (Lampetra) и некоторых лососевых рыб (Oncorhynchus), которые резко отличаются временем нереста; между особями каждой из рас существует высокая степень изоляции. Среди растений известны случаи генетически обусловленного сдвига в период цветения, создающего биологическую изоляцию этих форм — явление фенологического полиморфизма (рис. 9.6).
В природе обычна биотопическая изоляция, при которой потенциальные партнеры по спариванию встречаются, так как они реже предпочитают разные места обитания. Так, часть зябликов (Fringilla coelebs) гнездится в лесах таежного типа, а другая — в невысоких и редких насаждениях с большим числом полян. Потенциальная возможность перекрестного спаривания особей этих групп ограничена. Интересный пример биотопической изоляции — симпатрические внутривидовые формы у обыкновенной кукушки (Cuculus canorus). В Европе обитает несколько «биологических рас» кукушек, различающихся генетически закрепленной окраской яиц. В Восточной Европе одни откладывают голубые яйца в гнезда обыкновенной горихвостки и лугового чекана, другие — светлые в крапинку яйца — в гнезда мелких воробьиных птиц, имеющих яйца сходной окраски. Изоляция между этими формами кукушек поддерживается за счет уничтожения видами-хозяевами недостаточно замаскированных яиц. У многих видов предпочтение биотопа — эффективный изоляционный механизм.
Большое значение в возникновении и поддержании биологической изоляции у близких форм имеет этнологическая изоляция — осложнения спаривания, обусловленные особенностями поведенияВелико разнообразие способов этологической изоляции у животных. Ничтожные на первый взгляд отличия в ритуале ухаживания и обмене зрительными, звуковыми, химическими раздражителями будут препятствовать продолжению ухаживания. Возможно, у животных этологические механизмы — наиболее обширная группа из докопуляционных изолирующих механизмов.
На рис. 9.7 показан характер световых вспышек у североамериканских светляков рода Photurus. Различия между близкими видами, а иногда и разными группами популяций внутри вида, определяются длительностью, частотой и интенсивностью вспышек.
Важным изолирующим механизмом, затрудняющим скрещивание близких видов, оказывается возникновение морфофизиологических различий в органах размножения (морфофизиологическая изоляция), а также специализация в опылении у растений.
У животных близких видов различия ко-пулятивных органов особенно характерны для некоторых легочных моллюсков, насекомых, а среди млекопитающих — ряда групп грызунов.
Описанные главные механизмы изоляции обычно достаточны для предотвращения в природных условияхвозможности скрещивания форм, принадлежащих к разным видам, и понижения эффективности скрещивания у внутривидовых форм, далеко разошедшихся в процессе эволюции (подвидов, групп удаленных географических популяций и т. п.).
Рис. 9.7. Пример эколого-этологической изоляции. Характеристика световых вспышек североамериканских светляков рода Photurus. Высота и длина пятен в таблице указывают на интенсивность и длительность вспышки (из Э. Майра, 1968)
Вторая большая группа изолирующих механизмов в природе связана с возникновением изоляции после оплодотворения (собственно-генетическая изоляция), включающей гибель зигот после оплодотворения, развитие полностью или частично стерильных гибридов, а также пониженную жизнеспособность гибридов.
При межвидовом спаривании часто образуются вполне жизнеспособные гибриды, но у них, как правило, не развиваются нормальные половые клетки. В случае же нормального развития гамет гибриды оказываются малоплодовитыми. В природе есть случаи такой изоляции посредством гибридизации: на границе обитания двух близких форм постоянно существует зона, населенная вполне жизнеспособными. гибридными особями, но их потомство либо ослаблено и не выдерживает конкуренции с более сильными особями родительских видов, либо нежизнеспособно. На существование такого гибридизационного барьера между видами обычно указывает наличие в природе устойчивых гибридных зон между близкими видами. Такие гибридные зоны известны для некоторых насекомых, серой и черной ворон в Европе (рис. 9.8) и других форм.
Значение изоляции в эволюции. Изоляция как эволюционный фактор не создает новых генотипов или внутривидовых форм. Значение изоляции в процессе эволюции состоит в том, что она закрепляет и усиливает начальные стадии генотипической дифференцировки, а также в том, что разделенные барьерами части популяции или вида неизбежно попадают под различное давление отбора. Изоляция ведет к сохранению специфичности генофонда ди-вергирующих форм.
Оценивая роль трех элементарных эволюционных факторов, можно сказать, что первые два фактора (мутационный процесс и волны жизни)—факторы-поставщики элементарного эволюционного материала (хотя и действуют совершенно по-разному), а изоляция оказывается фактором-усилителем генетических различий между группами особей.
Механизм действия трех факторов различен, но общими для них являются ненаправленность, неопределенность и стохастичность действия. В природе три эволюционных фактора действуют совместно, однако роль каждого может усиливаться в зависимости от конкретной обстановки. Даже совместное действие трех элементарных факторов не обеспечивает устойчивого протекания направленного процесса эволюции.
Направленность эволюции придает действие естественного отбора.
14. Естественный отбор и механизмы его действия. Факторы, определяющие
эффективность и скорость ественного отбора.
Естественный отбор — процесс, изначально определённый Чарльзом Дарвином как приводящий к выживанию и преимущественному размножению более приспособленных к данным условиям среды особей, обладающих полезными наследственными признаками. В соответствии с теорией Дарвина и современной синтетической теорией эволюции, основным материалом для естественного отбора служат случайные наследственные изменения — рекомбинация генотипов, мутации и их комбинации.
При отсутствии полового процесса естественный отбор приводит к увеличению доли данного генотипа в следующем поколении. Однако естественный отбор «слеп» в том смысле, что он «оценивает» не генотипы, а фенотипы, и преимущественная передача следующему поколению генов особи, обладающей полезными признаками, происходит независимо от того, являются ли эти признаки наследуемыми.
Более успешной считают особь, которая оставила большее число потомков (то есть копий своих генов).[1].
При наличии полового процесса представления о естественном отборе (ЕО) сильно усложняются и утрачивают изначальную определенность. Так, регулярный кроссинговер не позволяет говорить об увеличении доли того или иного генотипа в следующем поколении, поскольку в 2-полых популяциях индивидуальные генотипы уникальны и неповторимы. И из поколения в поколение они не переходят. Остается преимущественная передача очередному поколению генов особей, отличающихся сравнительно высокой «приспособленностью». То есть, по сути речь идет об отборе отдельных аллелей, а не целостных генотипов.
Однако и по отношению к такому (ослабленному) пониманию ЕО возникают серьёзные вопросы. Как известно, диплоидные генотипы несут двойной набор генетической информации, причем аллели в двух гомологичных хромосомах часто разные. На фенотипе особи рецессивные аллели часто не сказываются. В силу кроссинговера и случайных сочетаний гаплоидных гамет при формировании зиготы в генотипах потомства оказываются далеко не все аллели, которые ответственны за формирование высоко «приспособленных» родительских фенотипов. И наоборот, передаются многие из тех аллелей, которые не проявились в фенотипах родителей.
Иначе говоря, из поколение в поколение переходят многие гены вовсе не из числа «лучших» — ни в каком смысле. И наоборот: передаются далеко не все «лучшие»; случайности кроссинговера и рекомбинации. Таким образом, своеобразный механизм наследования в 2-полых популяциях ставит под сомнение традиционные представления об отборе лучше «приспособленных» и придает действию ЕО иной характер. В сущности имеет место более-менее последовательная выбраковка «худших» (ущербных) аллелей — и только. Хотя и столь «нечеткого» действия ЕО достаточно для достижения той «приспособленности» к условиям обитания, какая наблюдается в живой природе. Механизм естественного отбора
Мысль о том, что виды могут изменяться под действием отбора, неоднократно высказывали разные учёные начиная с античных времён, в том числе некоторые английские авторы начала XIX века. Однако широкое признание концепция естественного отбора получила после того, как в 1858 году английские учёные Чарльз Дарвин и Альфред Уоллес изложили в своих статьях, опубликованных в одном и том же номере журнала Journal of the Proceedings of the Linnean Society of London. Zoology, идею о том, что в живой природе действует механизм, подобный искусственному отбору, и в особенности после выхода в свет в 1859 году книги Дарвина «Происхождение видов». Смысл их идеи состоит в том, что для появления удачных созданий природе вовсе не обязательно понимать и анализировать ситуацию, а можно действовать наугад. Достаточно создавать широкий спектр разнообразных особей — и, в конечном счёте, выживут наиболее приспособленные.
Сначала появляется особь с новыми, совершенно случайными, свойствами
Потом она оказывается или не оказывается способной оставить потомство, в зависимости от этих свойств
Наконец, если исход предыдущего этапа оказывается положительным, то она оставляет потомство и её потомки наследуют новоприобретённые свойства
В настоящее время отчасти наивные взгляды самого Дарвина оказались частично переработаны. Так, Дарвин представлял, что изменения должны происходить очень плавно, а спектр изменчивости является непрерывным. Сегодня, однако, механизмы естественного отбора объясняются при помощи генетики, которая вносит некоторое своеобразие в эту картину. Мутации в генах, которые работают на первом этапе описанного выше процесса, являются дискретными изменениями генотипа. Ясно, однако, что основной смысл идеи Дарвина остался без изменений.
[править]Формы естественного отбора
Существуют разные классификации форм отбора. Широко используется классификация, основанная на характере влияния форм отбора на изменчивость признака в популяции.
[править]Движущий отбор
Движущий отбор — форма естественного отбора, которая действует при направленном изменении условий внешней среды. Описали Дарвин и Уоллес. В этом случае особи с признаками, которые отклоняются в определённую сторону от среднего значения, получают преимущества. При этом иные вариации признака (его отклонения в противоположную сторону от среднего значения) подвергаются отрицательному отбору. В результате в популяции из поколения к поколению происходит сдвиг средней величины признака в определённом направлении. При этом давление движущего отбора должно отвечать приспособительным возможностям популяции и скорости мутационных изменений (в ином случае давление среды может привести к вымиранию).
Примером действия движущего отбора является «индустриальный меланизм» у насекомых. «Индустриальный меланизм» представляет собой резкое повышение доли меланистических (имеющих тёмную окраску) особей в тех популяциях насекомых (например, бабочек), которые обитают в промышленных районах. Из-за промышленного воздействия стволы деревьев значительно потемнели, а также погибли светлые лишайники, из-за чего светлые бабочки стали лучше видны для птиц, а тёмные — хуже. В XX веке в ряде районов доля тёмноокрашенных бабочек в некоторых хорошо изученных популяциях березовой пяденицы в Англии достигла 95 %, в то время как впервые тёмная бабочка (morfa carbonaria) была отловлена в 1848 году.
Движущий отбор осуществляется при изменении окружающей среды или приспособлении к новым условиям при расширении ареала. Он сохраняет наследственные изменения в определённом направлении, перемещая соответственно и норму реакции. Например, при освоении почвы как среды обитания у различных неродственных групп животных конечности превратились в роющие.
[править]Стабилизирующий отбор
Стабилизирующий отбор — форма естественного отбора, при которой его действие направлено против особей, имеющих крайние отклонения от средней нормы, в пользу особей со средней выраженностью признака. Понятие стабилизирующего отбора ввел в науку и проанализировал И. И. Шмальгаузен.
Описано множество примеров действия стабилизующего отбора в природе. Например, на первый взгляд кажется, что наибольший вклад в генофонд следующего поколения должны вносить особи с максимальной плодовитостью. Однако наблюдения над природными популяциями птиц и млекопитающих показывают, что это не так. Чем больше птенцов или детёнышей в гнезде, тем труднее их выкормить, тем каждый из них меньше и слабее. В результате наиболее приспособленными оказываются особи со средней плодовитостью.
Отбор в пользу средних значений был обнаружен по множеству признаков. У млекопитающих новорождённые с очень низким и очень высоким весом чаще погибают при рождении или в первые недели жизни, чем новорождённые со средним весом. Учёт размера крыльев у воробьёв, погибших после бури в 50-х годах под Ленинградом, показал, что большинство из них имели слишком маленькие или слишком большие крылья. И в этом случае наиболее приспособленными оказались средние особи.
[править]Дизруптивный отбор
Дизруптивный (разрывающий) отбор — форма естественного отбора, при котором условия благоприятствуют двум или нескольким крайним вариантам (направлениям) изменчивости, но не благоприятствуют промежуточному, среднему состоянию признака. В результате может появиться несколько новых форм из одной исходной. Дарвин описывал действие дизруптивного отбора, считая, что он лежит в основе дивергенции, хотя и не мог привести доказательств его существования в природе. Дизруптивный отбор способствует возникновению и поддержанию полиморфизма популяций, а в некоторых случаях может служить причиной видообразования.
Одна из возможных в природе ситуаций, в которой вступает в действие дизруптивный отбор, — когда полиморфная популяция занимает неоднородное местообитание. При этом разные формы приспосабливаются к различным экологическим нишам или субнишам.
Примером дизруптивного отбора является образование двух рас у погремка большого на сенокосных лугах. В нормальных условиях сроки цветения и созревания семян у этого растения покрывают всё лето. Но на сенокосных лугах семена дают преимущественно те растения, которые успевают отцвести и созреть либо до периода покоса, либо цветут в конце лета, после покоса. В результате образуются две расы погремка — ранне- и позднецветущая.
Дизруптивный отбор осуществлялся искусственно в экспериментах с дрозофилами. Отбор проводился по числу щетинок, оставлялись только особи с малым и большим количеством щетинок. В результате примерно с 30-го поколения две линии разошлись очень сильно, несмотря на то, что мухи продолжали скрещиваться между собой, осуществляя обмен генами. В ряде других экспериментов (с растениями) интенсивное скрещивание препятствовало эффективному действию дизруптивного отбора.
[править]Половой отбор
Половой отбор - это естественный отбор на успех в размножении. Выживание организмов является важным, но не единственным компонентом естественного отбора. Другим важнейшим компонентом является привлекательность для особей противоположного пола. Дарвин назвал это явление половым отбором. «Эта форма отбора определяется не борьбой за существование в отношениях органических существ между собою или с внешними условиями, но соперничеством между особями одного пола, обычно самцами, за обладание особями другого пола». Признаки, которые снижают жизнеспособность их носителей, могут возникать и распространяться, если преимущества, которые они дают в успехе размножения, значительно выше, чем их недостатки для выживания. Было предложено две основные гипотезы о механизмах полового отбора. Согласно гипотезе «хороших генов» самка «рассуждает» следующим образом: «Если этот самец, несмотря на его яркое оперение и длинный хвост, каким-то образом умудрился не погибнуть в лапах хищника и дожить до половой зрелости, то, следовательно, он обладает хорошими генами, которые позволили ему это сделать. Значит, его стоит выбрать в качестве отца для своих детей: он передаст им свои хорошие гены». Выбирая ярких самцов, самки выбирают хорошие гены для своих потомков. Согласно гипотезе «привлекательных сыновей» логика выбора самок несколько иная. Если яркие самцы, по каким бы то ни было причинам, являются привлекательными для самок, то стоит выбирать яркого отца для своих будущих сыновей, потому что его сыновья унаследуют гены яркой окраски и будут привлекательными для самок в следующем поколении. Таким образом, возникает положительная обратная связь, которая приводит к тому, что из поколения в поколение яркость оперения самцов все более и более усиливается. Процесс идет по нарастающей до тех пор, пока не достигнет предела жизнеспособности. В выборе самцов самки не более и не менее логичны, чем во всем остальном их поведении. Когда животное чувствует жажду, оно не рассуждает, что ему следует попить воды, для того чтобы восстановить водно-солевой баланс в организме — оно идет на водопой, потому что чувствует жажду. Точно так же и самки, выбирая ярких самцов, следуют своим инстинктами — им нравятся яркие хвосты. Все те, кому инстинкт подсказывал иное поведение, все они не оставили потомства. Таким образом, мы обсуждали не логику самок, а логику борьбы за существование и естественного отбора — слепого и автоматического процесса, который, действуя постоянно из поколения в поколение, сформировал все то удивительное разнообразие форм, окрасок и инстинктов, которое мы наблюдаем в мире живой природы.
[править]Положительный и отрицательный отбор
[править]Положительный отбор
Положительный отбор — форма естественного отбора. Его действие противоположно отсекающему отбору. Положительный отбор увеличивает в популяции число особей, обладающих полезными признаками, повышающими жизнеспособность вида в целом.
[править]Отсекающий (отрицательный) отбор
Отсекающий отбор — форма естественного отбора. Его действие противоположно положительному отбору. Отсекающий отбор выбраковывает из популяции подавляющее большинство особей, несущих признаки, резко снижающие жизнеспособность при данных условиях среды. С помощью отсекающего отбора из популяции удаляются сильно вредные аллели. Также отсекающему отбору могут подвергаться особи с хромосомными перестройками и набором хромосом, резко нарушающими нормальную работу генетического аппарата.
[править]Роль естественного отбора в эволюции
Чарльз Дарвин полагал естественный отбор основополагающим фактором эволюции живого (селекционизм в биологии). Накопление в конце XIX — начале XX века сведений по генетике, в частности обнаружение дискретного характера наследования фенотипических признаков, подтолкнуло многих исследователей к пересмотру указанного тезиса Дарвина: в качестве чрезвычайно важных факторов эволюции стали рассматриваться мутации генотипа (мутационизм Гуго де Фриза, сальтационизм Рихарда Гольдшмитда и др.). С другой стороны, открытие известных корреляций среди признаков родственных видов (закон гомологических рядов) Н. И. Вавилова привело к формулировке гипотез об эволюции на основе закономерностей, а не случайной изменчивости (номогенез Л. С. Берга, батмогенез Эдварда Дринкера Копа и других). В 1920-1940-е годы в эволюционной биологии интерес к селекционистским теориям возродился благодаря синтезу классической генетики и теории естественного отбора. Разработанная в результате этого синтетическая теория эволюции, часто называемая неодарвинизмом, опирается на количественный анализ частоты аллелей в популяциях, изменяющейся под влиянием естественного отбора. Тем не менее, открытия последних десятилетий в различных областях научного знания — от молекулярной биологии с её теорией нейтральных мутаций Мотоо Кимуры и палеонтологии с её теорией прерывистого равновесия Стивена Джея Гоулда и Найлза Элдриджа (в которой вид понимается как относительно статическая фаза эволюционного процесса) до математики с её теорией бифуркаций и фазовых переходов — свидетельствуют о недостаточности классической синтетической теории эволюции для адекватного описания всех аспектов биологической эволюции. Дискуссия о роли различных факторов в эволюции продолжается и сегодня, и эволюционная биология подошла к необходимости своего очередного, третьего синтеза.
[править]Интересные факты
Дарвин долго не решался обнародовать свою теорию, так как видел проблему муравьёв[2], которую можно было объяснить только с позиций генетики.
15. Основные формы естественного отбора.
16. Творческая роль естественного отбора.
17. Адаптации как результат действия естественного отбора.
Возникновение адаптаций в результате естественного отбора
Адаптациями называются свойства и признаки организмов, которые обеспечивают приспособление к той среде, в которой эти организмы живут. Адаптацией также называют процесс возникновения приспособлений. Выше мы рассмотрели, как некоторые адаптации возникают в результате естественного отбора. Популяции березовой пяденицы приспособились к изменившимся внешним условиям благодаря накоплению мутаций темной окраски. В популяциях человека, населяющих малярийные районы, адаптация возникла благодаря распространению мутации серповидно-клеточной анемии. И в том, и в другом случае, адаптация достигается за счет действия естественного отбора.
При этом материалом для отбора служит наследственная изменчивость, накопленная в популяциях. Поскольку разные популяции отличаются друг от друга по набору накопленных мутаций, то к одним и тем же факторам внешней среды они приспосабливаются по-разному. Так, африканские популяции адаптировались к жизни в малярийных районах за счет накопления мутаций серповидно-клеточной анемии HbS, а в популяциях, населяющих юго-восточную Азию устойчивость к малярии сформировалась на основе накопления ряда других мутаций, которые в гомозиготном состоянии также вызывают болезни крови, а в гетерозиготном - обеспечивают защиту от малярии.
Эти примеры иллюстрируют роль естественного отбора в формировании адаптаций. Нужно, однако, ясно понимать, что это – частные случаи относительно простых адаптаций, возникающих за счет селективного размножения носителей единичных «полезных» мутаций. Маловероятно, что большинство адаптаций возникло таким путем.
Покровительственная, предостерегающая и подражательная окраска. Рассмотрим, например, такие широко распространенные адаптации, как покровительственная, предостерегающая и подражательная окраска (мимикрия). Покровительственная окраска позволяет животным становится незаметными, сливаясь с субстратом. Одни насекомые поразительно сходны с листьями деревьев, на которых они обитают, другие напоминают засохшие веточки или шипы на стволах деревьев. Эти морфологические адаптации дополняются поведенческими приспособлениями. Насекомые выбирают для укрытия именно те места, где они менее заметны.
Несъедобные насекомые и ядовитые животные - змеи и лягушки, имеют яркую, предостерегающую окраску. Хищник, раз столкнувшись с таким животным, надолго ассоциирует этот тип окраски с опасностью. Этим пользуются некоторые неядовитые животные. Они приобретают поразительное сходство с ядовитыми, и тем самым снижают опасность со стороны хищников. Уж имитирует окраску гадюки, муха подражает пчеле. Это явление называется мимикрией.
Как возникли все эти удивительные приспособления? Маловероятно, чтобы единичная мутация могла обеспечивать такое точное соответствие между крылом насекомого и живым листом, между мухой и пчелой. Невероятно, чтобы единственная мутация заставляла покровительственно окрашенное насекомое прятаться именно на тех листьях, на которые оно похоже. Очевидно, что такие приспособления как покровительственная и предостерегающая окраски и мимикрия возникали путем постепенного отбора всех тех мелких уклонений в форме тела, в распределении определенных пигментов, во врожденном поведении, которые существовали в популяциях предков этих животных. Одной из важнейших характеристик естественного отбора является его кумулятивность – его способность накапливать и усиливать эти уклонения в ряду поколений, слагая изменения отдельных генов и контролируемых ими систем организмов.
Самая интересная и трудная проблема – это начальные стадии возникновения адаптаций. Понятно, какие преимущества дает почти идеальное сходство богомола с сухим сучком. Но какие преимущества могли быть у его далекого предка, который лишь отдаленно напоминал сучок? Неужели хищники так глупы, что их можно так легко обмануть? Нет, хищники отнюдь не глупы, и естественные отбор из поколения в поколение «учит» их все лучше и лучше распознавать уловки их добычи. Даже идеальное сходство современного богомола с сучком не дает ему 100-процентной гарантии, что ни одна птица его никогда не заметит. Однако его шансы ускользнуть от хищника выше, чем у насекомого с менее совершенной покровительственной окраски. Точно также, у его далекого предка, лишь чуть-чуть похожего на сучок, шансы на жизнь были несколько выше, чем у его родственника вовсе на сучок не похожего. Конечно, птица, которая сидит рядом с ним, в ясный день легко его заметит. Но если день туманный, если птица не сидит рядом, а пролетает мимо и решает не тратить времени на то, что может быть богомолом, а может быть и сучком, тогда и минимальное сходство сохраняет жизнь носителю этого едва заметного сходства. Его потомки, которые унаследуют это минимальное сходство, будут более многочисленны. Их доля в популяции станет больше. Это осложнит жизнь птицам. Среди них станут более успешными те, кто точнее будет распознавать замаскированную добычу. Вступает в действие тот самый принцип Красной Королевы, который мы обсуждали в параграфе, посвященном борьбе за существование. Чтобы сохранить преимущество в борьбе за жизнь, достигнутое за счет минимального сходства, виду-жертве приходится меняться.
Естественный отбор подхватывает все те мельчайшие изменения, которые усиливают сходство в окраске и форме с субстратом, сходство между съедобным видом и тем несъедобным видом, которому он подражает. Следует учитывать, что разные виды хищников пользуются разными методами поиска добычи. Одни обращают внимание на форму, другие на окраску, одни обладают цветным зрением, другие нет. Поэтому естественный отбор автоматически усиливает, насколько это возможно, сходство между имитатором и моделью и приводит к тем изумительным адаптациям, которые мы наблюдаем в живой природе.
Возникновение сложных адаптаций. Многие адаптации производят впечатление тщательно продуманных и направленно спланированных устройств. Как такая сложнейшая структура как глаз человека могла возникнуть путем естественного отбора случайно возникавших мутаций?
Ученые предполагают, что эволюция глаза началась с небольших групп светочувствительных клеток на поверхности тела наших очень далеких предков, живших около 550 млн. лет назад. Способность различать свет и тьму была, безусловно, полезна для них, повышала их шансы на жизнь по сравнению с их абсолютно слепыми сородичами. Случайно возникшее искривление «зрительной» поверхности улучшило зрение, это позволяло определить направление на источник света. Возник глазной бокал. Вновь возникающие мутации могли вести к сужению и расширению отверстия глазного бокала. Сужение постепенно улучшало зрение – свет стал проходить через узкую диафрагму. Как видите, каждый шаг повышал приспособленность тех особей, которые менялись в «правильном» направлении. Светочувствительные клетки формировали сетчатку. Со временем в передней части глазного яблока сформировался хрусталик, выполняющий функцию линзы. Он возник, по-видимому, как прозрачная двухслойная структура, наполненная жидкостью.
Ученые попытались смоделировать этот процесс на компьютере. Они показали, что глаз, подобный сложному глазу моллюска, мог возникнуть из слоя фоточувствительных клеток при относительно мягком отборе всего за 364 000 поколений. Иными словами, животные, у которых смена поколений происходит каждый год, могли сформировать полностью развитый и оптически совершенный глаз в менее, чем за полмиллиона лет. Эта очень короткий срок для эволюции, если учесть что средний возраст вида у моллюсков равняется нескольким миллионам лет.
Все предполагаемые стадии эволюции глаза человека мы можем обнаружить среди ныне живущих животных. Эволюция глаза шла разными путями в разных типах животных. Благодаря естественному отбору независимо возникло множество разных форм глаза, и человеческий глаз – только один из них, причем не самый совершенный
Если мы внимательно рассмотрим конструкцию глаза человека и других позвоночных животных, мы обнаружим целый ряд странных несообразностей. Когда свет попадает в глаз человека он проходит через хрусталик и попадает на светочувствительные клетки сетчатки. Свет вынужден пробиваться через густую сеть капилляров и нейронов, чтобы попасть на фоторецепторный слой. Как это ни удивительно, но нервные окончания подходят к светочувствительным клеткам не сзади, а спереди! Более того, нервные окончания собираются в оптический нерв, который отходит от центра сетчатки, и создает тем самым слепое пятно. Чтобы компенсировать затенение фоторецепторов нейронами и капиллярами и избавится от слепого пятна, наш глаз постоянно движется, посылая в мозг серию разных проекций одного и того же изображения. Наш мозг производит сложнейшие операции, складывая эти изображения, вычитая тени, и вычисляя реальную картину. Всех этих сложностей можно было бы избежать, если бы нервные окончания подходили к нейронам не спереди, а сзади как, например, у осьминога.
Само несовершенство глаза позвоночных проливает свет на механизмы эволюции путем естественного отбора. Мы уже не раз говорили о том, что отбор всегда действует «здесь и сейчас». Он сортирует разные варианты уже существующих структур, выбирая и слагая вместе лучшие из них: лучшие «здесь и сейчас», безотносительно к тому, во что эти структуры могут превратиться в далеком будущем. Поэтому ключ к объяснению и совершенств и несовершенств современных структур следует искать в прошлом. Ученые считают, что все современные позвоночные произошли от животных подобных ланцетнику. У ланцетника светочувствительные нейроны располагаются на переднем конце нервной трубки. Перед ними расположены нервные и пигментные клетки, которые прикрывают фоторецепторы от света попадающего спереди. Ланцетник принимает световые сигналы, приходящие с боков его прозрачного тела. Можно думать, что у общего предка позвоночных глаз был устроен сходным образом. Затем эта плоская структура стала преобразовываться в глазной бокал. Передняя часть нервной трубки впячивалась внутрь, и нейроны, находившиеся впереди рецепторных клеток, оказались поверх них. Процесс развития глаза у эмбрионов современных позвоночных в известном смысле воспроизводит последовательность событий, происходивших в далеком прошлом.
Эволюция не создает новых конструкций «с чистого листа», она меняет (часто неузнаваемо меняет) старые конструкции, так чтобы каждый этап этих изменений был приспособительным. Любое изменение должно повышать приспособленность его носителей или, хотя бы, не снижать ее. Эта особенность эволюции ведет к неуклонному совершенствованию различных структур. Она же является причиной несовершенства многих адаптаций, странных несообразностей в строении живых организмов.
Следует помнить, однако, что все приспособления, сколь бы совершенны они ни были, носят относительный характер. Понятно, что развитие способности к полету не очень хорошо совмещается с возможностью быстро бегать. Поэтому птицы, обладающие наилучшими способностями к полету, — плохие бегуны. Напротив, страусы, которые не способны летать, прекрасно бегают. Приспособление к определенным условиям может быть бесполезно или даже вредно при появлении новых условий. Однако условия обитания меняются постоянно и иногда очень резко. В этих случаях накопленные ранее адаптации могут затруднить формирование новых, что может вести к вымиранию больших групп организмов, как это случилось более 60—70 млн лет назад с некогда очень многочисленными и разнообразными динозаврами.
18. Маскирующие окраски и формы как эволюционные адаптации.
Адаптация — процесс приспособления, процесс возникновения признака, соответствующего условиям среды. Процесс возникновения приспособлений И. И. Шмальгаузен назваладаптациогенезом, а процесс приспособительного преобразования организмов —адаптациоморфозом. Георгиевский пришел к заключению, что "по своему содержаниюестественный отбор и адаптациогенез — понятия тождественные". Знак равенства ставил между ними и Ч. Дарвин, характеризуя отбор как "выживание наиболее приспособленных".
Термин "адаптация" имеет три важных смысла: приспособление, процесс приспособления, приспособленность.
Адаптация — приспособление. Это любая структура, любой признак, возникший в процессе эволюции, обеспечивающий успешное выживание и размножение. Особенности габитуса, внешнего строения. Крючочки на плодах лопуха — приспособление для распространения животными, покрытыми шерстью. Длинный мех — адаптация к холоду. Корни верблюжьей колючки уходят на глубину до 30 м — приспособление к жизни в пустыне. Ботрии и ботридии цестод — адаптация для прикрепления паразита к стенке кишечника хозяина. Аскарида имеет многослойную и сложную кутикулу Это приспособление к обитанию в среде, богатой ферментами.
Приспособительная окраска — покровительственная, предостерегающая, мимикрия. Покровительственная окраска — это окраска, делающая организмы менее заметными в местах их обитания.
Покровительственная окраска — это криптическая окраска, маскировка животных: обитатели пустыни имеют желто-бурую окраску тела; яйца птиц, гнездящихся в траве, имеют цвет, соответствующий фону; сезонная окраска зайца-беляка, песца, горностая, белой куропатки; серое оперение перепела; темная окраска верха и светлая окраска низа у рыб; расчленяющая окраска тигра и зебры.
Предостерегающая окраска - яркая, контрастная (красная с черным, черная с желтым) окраска у животных, имеющих ядовитые или пахучие железы. Предупреждающая окраска — это сигнал опасности для хищников о несъедобности данных организмов (осы, пчелы).
Мимикрия (греч. мимикос — уподобление, подражание) — подражательное сходство незащищенного организма с защищенным или несъедобным. Мимикрия бейтсовская — это сходство беззащитного съедобного с несъедобным ярко окрашенным. Мюллеровская мимикрия — сходство между двумя (и более) несъедобными видами (многие виды ос сходны по окраске и форме тела).
Миметизм - внешнее сходство незащищенных животных с растениями и предметами неживой природы (морской конек похож на водоросль, яйца кулика-сороки - на гальку);
Загар — адаптация к увеличению инсоляции. У дрожжей дупликация гена, контролирующего синтез фосфатазы, позволяет им жить в среде с более низкими концентрациями нужного субстрата. Богатые кремнием оболочки растений предохраняют их от поедания.
Физиологические адаптации — это функциональные адаптации (поддержание постоянной температуры, постоянства сахара в крови, выработка устойчивости к колебаниям солености, влажности). Скорость кровогока у эскимосов при охлаждении, в среднем, вдвое больше, чем у европейцев. Это приспособление позволяет эскимосам значительно легче, чем европейцам, сохранять тепловой баланс при охлаждении. Собака в покое в жаркую погоду увеличивает число дыхательных актов с 20 до 30 в минуту: частое дыхание - адаптация против жары. У кедрового стланника ветви с наступлением морозов полегают — это адаптация против сильных ветров и морозов.
Этологические адаптации — изменение поведения, направленное на выживание отдельных организмов.
19. Демонстрационные окраски и формы как эволюционные адаптации
В ходе эволюции добиваются успеха те организмы, которые лучше других приспособлены к окружающей среде. Механизмы эволюции обеспечивают соответствие развивающейся живой системы условиям её существования - высокую приспособленность жизненных форм к внешней среде. В основе приспособленности лежат процессы самовоспроизведения макромолекул и живых организмов и дискретность всего живого на Земле. У организмов в процессе эволюции выработалось множество разнообразных относительных приспособлений (адаптаций) к окружающей среде. Адаптацией считается любая особенность особи, популяции, вида или сообщества организмов, которая способствует успеху в конкуренции и обеспечивает устойчивость к абиотическим факторам. Это позволяет организмам существовать в данных условиях среды и оставлять потомство. Критериями адаптации являются: жизнеспособность, конкурентоспособность и фертильность.
Жизнеспособность - это способность организма жить и нормально развиваться в условиях среды.
Конкурентоспособность -- это способность организма добиваться успеха в борьбе за средства жизни.
Фертильность - это способность организмов нормально размножаться.
1. Виды адаптации
Все адаптации делят на аккомодации и эволюционные адаптации. Аккомодации представляют собой обратимый процесс. Они возникают при резком изменении условий среды. Например, при переселении животные попадают в новую для них обстановку, но постепенно привыкают к ней. Например, человек, переселившийся из средней полосы в тропики или на Крайний Север, некоторое время испытывает дискомфорт, но со временем привыкает к новым условиям. Эволюционная адаптация необратима и возникшие изменения генетически закрепляются. Сюда относят все приспособления, на которые действует естественный отбор. Например, покровительственная окраска или быстрый бег.
Приспособления также делят на организменные и видовые. Организменные адаптации в свою очередь подразделяются на морфологические, физиологические, биохимические и этологические.
Морфологические адаптации проявляются в преимуществах строения, покровительственной окраске, предостерегающей окраске, мимикрии, маскировке, приспособительном поведении.
Преимущества строения - это оптимальные пропорции тела, расположение и густота волосяного или перьевого покрова и т.п. Хорошо известен облик водного млекопитающего - дельфина. Его движения легки и точны. Самостоятельная скорость движения в воде достигает 40 километров в час. Плотность воды в 800 раз выше плотности воздуха. Как дельфину удается преодолеть её? Помимо других особенностей строения идеальной приспособленности дельфина к среде обитания и образу жизни способствует форма тела. Торпедовидная форма тела позволяет избежать образования завихрений потоков воды, обтекающих дельфина.
Обтекаемая форма тела способствует быстрому передвижению животных и в воздушной среде. Маховые и контурные перья, покрывающие тело птицы, полностью сглаживают его форму. Птицы лишены выступающих ушных раковин, в полёте они обычно втягивают ноги. В результате птицы по скорости передвижения намного превосходят всех других животных. Например, сокол сапсан пикирует на свою жертву со скоростью до 290 километров в час. Птицы быстро двигаются даже в воде. Наблюдали антарктического пингвина, плывущего под водой со скоростью около 35 километров в час.
У животных, ведущих скрытный, затаивающийся образ жизни, полезным оказываются приспособления, придающие им сходство с предметами окружающей среды. Причудливая форма тела у рыб, обитающих в зарослях водорослей (морской конёк-тряпичник, рыба-клоун, морская игла и др.), помогает им успешно скрываться от врагов. Сходство с предметами среды обитания широко распространено у насекомых. Известны жуки, своим внешним видом напоминающие лишайники, цикады, сходные с шипами тех кустарников, среди которых они живут. Насекомые палочники похожи на небольшую бурую или зелёную веточку, а прямокрылые насекомые имитируют лист. Плоское тело имеют рыбы, ведущие придонный образ жизни (напр., камбала).
Покровительственная окраска позволяет быть незаметным среди окружающего фона. Благодаря покровительственной окраске организм становится трудно различимым и, следовательно, защищенным от хищников. Яйца птиц, откладываемые на песок или на землю, имеют серый и бурый цвет с пятнышками, сходный с цветом окружающей почвы. В тех случаях, когда яйца недоступны для хищников, они обычно лишены окраски. Гусеницы бабочек часто зелёные, под цвет листьев, или тёмные, под цвет коры или земли. Донные рыбы обычно окрашены под цвет песчаного дна (скаты и камбалы). При этом камбалы обладают ещё способностью менять окраску в зависимости от цвета окружающего фона. Способность менять окраску путём перераспределения пигмента в покровах тела известна и у наземных животных (хамелеон). Животные пустынь, как правило, имеют желто-бурую или песочно-желтую окраску. Однотонная покровительственная окраска свойственна как насекомым (саранча) и мелким ящерицам, так и крупным копытным (антилопы) и хищникам (лев).
Если фон среды не остается постоянным в зависимости от сезона года, многие животные меняют окраску. Например, обитатели средних и высоких широт (песец, заяц, горностай, белая куропатка) зимой имеют белую окраску, что делает их незаметными на снегу.
Вариант покровительственной окраски - расчленяющая окраска в виде чередования на теле светлых и тёмных полос и пятен. Зебры и тигр плохо видны уже на расстоянии 40-50 метров из-за совпадения полос на теле с чередованием света и тени в окружающей местности. Расчленяющая окраска нарушает представления о контурах тела.
Предостерегающая (угрожающая) окраска предупреждает потенциального врага о наличии защитных механизмов (наличие ядовитых веществ или специальных органов защиты). Предостерегающая окраска выделяет из окружающей среды яркими пятнами или полосами ядовитых, жалящих животных и насекомых (змеи, осы, шмели).
Эффективность предостерегающей окраски послужила причиной очень интересного явления - подражания (мимикрии). Мимикрией называется сходство в окраске, форме тела безопасных животных с ядовитыми и опасными животными. Отдельные виды мух, не имеющие жала, похожи на жалящих шмелей и ос, неядовитые змеи - на ядовитых. Во всех случаях сходство чисто внешнее и направлено на формирование определенного зрительного впечатления у потенциальных врагов. Сейчас известны два основных вида мимикрии: мимикрия Бейтса и мимикрия Мюллера.
При мимикрии Бейтса модель хорошо защищена и обычно имеет яркую, предостерегающую окраску. При мимикрии Мюллера сходными оказываются два и более несъедобных вида: в результате их сходства хищник скорее отучается схватывать таких животных. Первый тип мимикрии можно сравнить с мелкой фирмой, подделывающейся под рекламу какой-то хорошо известной крупной фирмы. Второй тип сравним с несколькими фирмами, которые для экономии средств пользуются общей рекламой. Пример мимикрии Бейтса: часто под видом ос скрываются беззащитные мухи, формой тела и жёлто-чёрной окраской имитирующие ос (муха-сирфида и муха-большеголовка). Пример мимикрии Мюллера: некоторые виды бабочек капустных белянок похожи на несъедобных южноамериканских геликонид.
Мимикрия - это результат гомологичных (одинаковых) мутаций у разных видов, которые помогают выжить незащищённым животным. Для видов-подражателей важно, чтобы их численность была невелика по сравнению с моделью, которой они подражают, иначе у врагов не будет выработан устойчивый отрицательный рефлекс на предостерегающую окраску. Низкая численность мимикрирующих видов поддерживается высокой концентрацией летальных генов в генофонде. В гомозиготном состоянии эти гены вызывают летальные мутации, в результате чего высокий процент особей не доживает до половозрелого состояния.
Кроме защитной окраски, у животных и растений наблюдаются и другие средства защиты. У растений нередко образуются иглы и колючки, защищающие их от поедания травоядными животными (кактусы, шиповник, боярышник, облепиха и др.). Такую же роль играют ядовитые вещества, обжигающие волоски, например у крапивы. Кристаллы щавелевокислого кальция, накапливающиеся в шипах некоторых растений, предохраняют их от поедания гусеницами, улитками и даже грызунами. Образования в виде твёрдого хитинового покрова у членистоногих (жуки, крабы), раковин у моллюсков, чешуи у крокодилов, панциря у броненосцев и черепах хорошо защищают их от многих врагов. Этому же служат иглы у ежа и дикобраза. Все эти приспособления могли появиться лишь в результате естественного отбора, т.е. преимущественного выживания лучше защищенных особей.
Маскировка - приспособления, при которых форма тела и окраска животных сливаются с окружающими предметами. Например, в тропических лесах многие змеи неразличимы среди лиан, лохматый морской конёк похож на водоросль, насекомые на коре деревьев похожи на лишайники (жуки, усачи, пауки, бабочки). Иногда приспособление к цвету и узору субстрата может осуществляться путём физиологического изменения окраски тела (каракатицы, скаты, камбалы, квакши) или переменой окраски при очередной линьке (кузнечики).
Защитное действие покровительственной окраски или формы тела повышается при сочетании её с соответствующим поведением. Приспособительное поведение - принятие определённых поз покоя (гусеницы некоторых насекомых в неподвижном состоянии очень похожи на сучок дерева; бабочка каллима со сложенными крыльями удивительно напоминает сухой лист дерева), либо, наоборот, демонстративное поведение, отпугивающее хищников. Помимо затаивания или демонстративного, отпугивающего поведения при приближении врага существует много других вариантов приспособительного поведения, обеспечивающего выживаемость взрослых особей или молоди. Сюда относится запасание корма на неблагоприятный сезон года. Особенно это присуще грызунам. Например, полёвка-экономка, распространённая в таёжной зоне, собирает зерна злаков, сухую траву, корешки - всего до 10 килограммов. Роющие грызуны (слепыши и др.) накапливают кусочки корней дуба, желуди, картофель, степной горошек - до 14 килограммов. Большая песчанка, живущая в пустынях Средней Азии, в начале лета срезает траву и затаскивает её в норы или оставляет на поверхности в виде стожков. Корм этот используется во второй половине лета, осенью и зимой. Речной бобр собирает обрубки деревьев, веток и пр., которые складывает в воду возле своего жилища. Склады эти могут достигать объёма 20 кубических метров. Запасы кормов делают и хищные животные. Норка и некоторые хорьки запасают лягушек, ужей, мелких зверьков и т.д. Примером приспособительного поведения служит и время наибольшей активности. В пустынях многие животные выходят на охоту ночью, когда спадает зной.
Физиологические адаптации - приобретение специфических особенностей обмена веществ в разных условиях среды. Они обеспечивают функциональные преимущества организма. Их условно разделяют на статические (постоянные физиологические параметры -- температура, водно-солевой баланс, концентрация сахара и т. п.) и динамические (адаптации к колебаниям действия фактора -- изменение температуры, влажности, освещенности, магнитного поля и т. п.).
Соответствующая форма и окраска тела, целесообразное поведение обеспечивают успех в борьбе за существование только тогда, когда эти признаки сочетаются с приспособленностью процессов жизнедеятельности к условиям обитания, т.е. с физиологической адаптацией. Без такой адаптации невозможно поддержание устойчивого обмена веществ в организме в постоянно колеблющихся условиях внешней среды. Приведем некоторые примеры.
Растения, живущие в полупустынных и пустынных районах, имеют многочисленные и разнообразные адаптации. Это и уходящий на десятки метров в глубь земли корень, извлекающий воду, и резкое уменьшение испарения воды благодаря особому строению кутикулы на листьях, и полная утрата листьев. У кактусов это преобразование особенно удивительно: преобразование стебля не только в орган, выполняющий опорную и проводящую функции, но и в структуру, запасающую воду и обеспечивающую фотосинтез. Крупные экземпляры кактусов накапливают до 2000 литров воды. Расходуется она медленно, так как клеточный сок содержит с органическими кислотами и сахарами также слизистые вещества, обладающие водоудерживающими свойствами. Стебли опунции даже после трёхмесячной засухи содержали почти 81% воды. Испарение воды значительно уменьшено благодаря ребристой структуре стеблей у кактусов, равномерно распределяющей свет и тень. Этому же способствует утолщение стенок эпидермиса, обычно покрытых слоем воска, наличие многочисленных колючек и волосков и многое другое.
У наземных амфибий большое количество воды теряется через кожу. Однако многие их виды проникают даже в пустыни и полупустыни. Выживание амфибий в условиях недостатка влаги в этих местах обитания обеспечивается целым рядом приспособлений. У них меняется характер активности: она приурочивается к периодам повышенной влажности. В умеренной зоне жабы и лягушки активны ночью и после выпадения дождей. В пустынях лягушки охотятся только ночью, когда влага конденсируется на почве и на растительности, а днем укрываются в норах грызунов. У пустынных видов амфибий, размножающихся во временных водоемах, личинки развиваются очень быстро и в сжатые сроки совершают метаморфоз.
Разнообразные механизмы физиологической адаптации к неблагоприятным условиям выработали птицы и млекопитающие. Многие пустынные животные перед наступлением засушливого сезона накапливают много жира: при его окислении образуется большое количество воды. Птицы и млекопитающие способны регулировать потери воды с поверхности дыхательных путей. Например, верблюд при лишении воды резко сокращает испарение как с дыхательных путей, так и через потовые железы.
У человека плохо регулируется солевой обмен, и поэтому он не может долго обходиться без пресной воды. Но рептилии и птицы, проводящие большую часть жизни в морских просторах и пьющие морскую воду, приобрели специальные железы, позволяющие им быстро избавляться от избытка солей.
Очень интересны приспособления, развивающиеся у ныряющих животных. Многие из них могут сравнительно долго обходиться без доступа кислорода. Например, тюлени ныряют на глубину 100--200 и даже 600 метров и находятся под водой 40--60 минут. Что позволяет ластоногим нырять на столь длительный срок? Это, прежде всего, большое количество особого пигмента, находящегося в мышцах,-- миоглобина. Миоглобин способен связать в 10 раз больше кислорода, чем гемоглобин. Кроме того, в воде целый ряд приспособлений обеспечивает гораздо более экономное расходование кислорода, чем при дыхании на поверхности.
Путем естественного отбора возникают и совершенствуются приспособления, облегчающие поиск пищи или партнера для размножения. Поразительно чувствительны органы химического чувства насекомых. Самцов непарного шелкопряда привлекает запах ароматической железы самки с расстояния 3 километров. У некоторых бабочек чувствительность рецепторов вкуса в 1000 раз превосходит чувствительность рецепторов человеческого языка. Ночные хищники, например совы, превосходно видят в условиях слабого освещения. У некоторых змей хорошо развита способность к термолокации. Они различают на расстоянии объекты, если разница их температур составляет всего 0,2 °С. Многие животные прекрасно ориентируются в пространстве с помощью эхолокации (летучие мыши, совы, дельфины).
Биохимические адаптации обеспечивают оптимальное течение биохимических реакций в клетке, например, упорядочение ферментативного катализа, специфическое связывание газов дыхательными пигментами, синтез нужных веществ в определенных условиях и т. п.
Этологические адаптации представляют собой все поведенческие реакции, направленные на выживание отдельных особей и, следовательно, вида в целом. Такими реакциями являются:
- поведение при поиске пищи и полового партнера,
- спаривание,
- выкармливание потомства,
- избегание опасности и защита жизни в случае угрозы,
- агрессия и угрожающие позы,
- незлобивость и многие другие.
Некоторые поведенческие реакции наследуются (инстинкты), другие приобретаются в течение жизни (условные рефлексы). У различных организмов соотношение инстинктивного и условнорефлекторного поведения неодинаково. Например, у беспозвоночных и низших хордовых преобладает инстинктивное поведение, а у высших млекопитающих (приматов, хищных) - условнорефлекторное. Высший уровень поведенческой адаптивности, основанный на механизмах высшей нервной деятельности, имеется у человека.
Особенно большое значение имеют приспособления, обеспечивающие защиту потомства от врагов.
Забота о потомстве может проявляться в разной форме. Многие рыбы охраняют икру, откладываемую между камнями, активно отгоняя и кусая приближающихся возможных врагов. Азовские и каспийские бычки откладывают икру в ямки, вырытые в дне, и охраняют её затем в течение всего развития. Самец колюшки строит гнездо с выходом и входом. Некоторые американские сомы прилепляют икру на брюхо и носят её на себе все время развития. Многие рыбы вынашивают икру во рту или даже в желудке. В это время родитель ничего не ест. Вылупившиеся мальки некоторое время держатся вблизи самки (или самца, в зависимости от вида) и при опасности прячутся в рот родителя. Существуют виды лягушек, у которых икринки развиваются в специальной выводковой сумке на спине или в голосовых мешках самца.
Наибольшая безопасность потомства достигается, очевидно, в тех случаях, когда зародыши развиваются в теле матери. Плодовитость в этих случаях снижается, однако это компенсируется возрастанием выживаемости молоди.
У членистоногих и низших позвоночных образующиеся личинки ведут самостоятельный образ жизни и не зависят от родителей. Но в некоторых случаях забота родителей о потомках проявляется в форме обеспечения их пищей. Знаменитый французский естествоиспытатель Ж.А.Фабр впервые описал такое поведение у одиночных ос. Осы нападают на жуков, пауков, сверчков, богомолов, гусениц различных бабочек, обездвиживают их, погружая жало точно в нервные узлы, и откладывают на них яйца. Вылупляющиеся личинки ос обеспечены пищей: они питаются тканями живой жертвы, растут и затем окукливаются.
Описанные примеры заботы о потомстве у членистоногих и низших позвоночных встречаются у очень небольшого числа видов. В большинстве случаев оплодотворённые яйца бывают брошены на произвол судьбы. Именно этим объясняется очень высокая плодовитость беспозвоночных и низших позвоночных животных. Большое число потомков в условиях высокой истребляемости молоди служит средством борьбы за существование вида.
Значительно более сложные и многообразные формы заботы о потомстве наблюдаются у высших позвоночных. Сложные инстинкты и способность к индивидуальному обучению позволяют им со значительно большим успехом выращивать потомство. Так, птицы откладывают оплодотворённые яйца в специальные сооружения - гнёзда, а не просто в наружную среду, как поступают все виды нижестоящих классов. Яйца развиваются под влиянием тепла, сообщаемого им телом родителей, и не зависят от случайностей погоды. Гнездо родители защищают от врагов теми или иными способами. Выведенных птенцов не оставляют на произвол судьбы, а длительное время выкармливают и охраняют их. Всё это резко повышает эффективность размножения у птиц.
Наивысшей степени развития достигают формы поведения у млекопитающих животных. Это проявляется и в отношении к детёнышам. Звери не только кормят своё потомство, но и обучают ловить добычу. Ещё Ч.Дарвин отмечал, что хищные звери учат своих детёнышей избегать опасностей, в том числе охотников.
Таким образом, особи с более совершенными формами заботы о потомстве выживают в большем числе и передают эти черты далее по наследству.
Видовые адаптации обнаруживаются при анализе группы особей одного вида, по своему проявлению они весьма разнообразны. Основными из них являются различные конгруэнции, уровень мутабильности, внутривидовой полиморфизм, уровень численности и оптимальная плотность населения.
Конгруэнции представляют собой все морфофизиологические и поведенческие особенности, которые способствуют существованию вида как целостной системы. Репродуктивные конгруэнции обеспечивают размножение. Некоторые из них непосредственно связаны с репродукцией (соответствие половых органов, приспособления к вскармливанию и др.), тогда как другие лишь опосредованно (различные сигнальные признаки: зрительные - брачный наряд, ритуальное поведение; звуковые - пение птиц, рев самца оленя во время гона и др.; химическими - различные аттрактанты, например, феромоны насекомых, выделения у парнокопытных, кошачьих, собачьих и др.).
К конгруэнциям относят все формы внутривидовой кооперации,-- конституциональной, трофической и репродуктивной. Конституциональная кооперация выражается в согласованных действиях организмов в неблагоприятных условиях, которые повышают шансы на выживание. Зимой пчелы собираются в шар, и выделяемое ими тепло расходуется на совместное согревание. При этом самая высокая температура будет в центре шара и особи с периферии (где холоднее) будут постоянно стремиться туда. Таким образом происходит постоянное перемещение насекомых и они совместными усилиями благополучно перезимуют. Также сбиваются в тесную группу пингвины во время насиживания, овцы в холодное время и др.
Трофическая кооперация состоит в объединении организмов с целью добывания пищи. Совместная деятельность в этом направлении делает процесс более продуктивным. Например, стая волков гораздо эффективнее охотится, нежели отдельная особь. При этом у многих видов имеет место разделение обязанностей - одни особи отделяют выбранную жертву от основного стада и гонят ее в засаду, где затаились их сородичи и т. д. У растений подобная кооперация выражается в совместном затенении почвы, что способствует удержанию в ней влаги.
Репродуктивная кооперация повышает успешность размножения и способствует выживанию потомства. У многих птиц особи собираются на токовищах, и в таких условиях облегчается поиск потенциального партнера. То же самое происходит на нерестилищах, лежбищах ластоногих и др. Вероятность опыления у растений повышается, когда они растут группами и расстояние между отдельными особями невелико.
Мутабильность - представляет собой частоту возникновения мутаций в единицу времени (количество поколений) и на один ген. Для каждого вида характерна своя частота, которая определяется уровнем стабильности генетического материала и устойчивостью к мутагенам. Мутации делают популяции гетероморфными и дают материал для осуществления отбора. Для вида опасна как чрезмерно высокая, так и недостаточная мутабильность. В первом случае возникает угроза целостности вида, а во втором -- невозможно осуществляться отбору.
Внутривидовой полиморфизм обусловливает уникальность сочетания аллелей у разных особей. Причиной полиморфизма служат половое размножение, которое обеспечивает комбинативную изменчивость, и мутации, изменяющие субстрат наследственности. Поддержание внутривидового полиморфизма обеспечивает устойчивость вида и гарантирует его существование в различных условиях среды.
Уровень численности определяет крайние значения количества особей вида. Снижение численности ниже порогового уровня ведет к гибели вида. Это связано с невозможностью встречи партнеров, нарушением внутривидовых адаптации и др. Чрезмерное увеличение численности также губительно, поскольку подрывает кормовую базу, способствует накоплению в популяции больных и ослабленных особей, у некоторых это приводит к развитию стресса.
Оптимальная плотность населения показывает специфические для каждого вида особенности сосуществования особей. Многие организмы предпочитают одиночный образ жизни и встречаются лишь для спаривания. Так ведут себя, например, тигры, леопарды, самцы слонов и др. У других сильно выражен инстинкт коллективности, поэтому они нуждаются в высокой численности. Например, самые многочисленные группы среди позвоночных образовывали американские странствующие голуби, стаи которых насчитывали миллиарды (!) особей. После того как человеком была подорвана их численность, странствующие голуби перестали размножаться и вид исчез.
Относительная целесообразность приспособлений (адаптаций)
Наличие разнообразных адаптаций к условиям жизни, зачастую экстремальным, - основа, позволяющая видам занять самые трудно доступные и неожиданные места обитания. Жизнь распространена повсеместно: от высоких широт Арктики, где обитают белые медведи, до охладительных контуров ядерных реакторов, где обитают некоторые виды микроорганизмов; от горных вершин, где встречаются лишайники, и до глубин океана, где обитают рыбы и другие животные. Каждый организм имеет множество различных адаптаций, но тем не менее представляет собой единое целое. Следовательно, у представителей любого вида осуществляется координация и взаимодействие адаптаций так, чтобы приспособленность организмов была максимальной.
Ни одна адаптация не является абсолютно идеальной. Некоторые из них достигают своего предела, например, глаз человека способен воспринимать отдельные фотоны (при соблюдении всех условий можно видеть пламя свечи на расстоянии двух км!), но в обычной жизни эта возможность не может быть достигнута, поскольку мешает атмосферная пыль, другие источники света, да и потребности такой у человека нет. Следовательно, такие адаптации (они называются абсолютными) не используются в полной мере. Однако большинство адаптации не достигает своего предельного значения (относительные адаптации).
Адаптации не бывают универсальными - каждая из них облегчает выполнение лишь определенной функции. Например, длинные крылья стрижа, позволяющие ему быстро летать, затрудняют взлет с ровной поверхности. Есть птицы, которые поедают ос и пчел, а также мух, мимикрирующих под них. Постоянный рост резцов у грызунов дает возможность грызть очень твердые предметы, однако если их не стачивать, отрастают так, что животное не может закрыть рот. Поэтому любой адаптивный признак оказывается целесообразным лишь в определенной среде. При резком изменении условий обитания чрезмерно развитые признаки могут оказаться нецелесообразными и принести вред организму. Поэтому после глобальных экологических катастроф преимущественно погибают высокоспециализированные виды (например, динозавры в меловом периоде палеозойской эры). То же самое происходит при отрицательном действии биотических факторов, например, гигантский торфяной олень был полностью уничтожен хищниками из-за своих огромных рогов, сделавших его малоподвижным.
Следует помнить, что все приспособления, сколь бы совершенными они не были, носят относительный характер. Дело в том, что естественный отбор есть результат переживания и более эффективного размножения в данном месте и в данное время. Значит, естественный отбор конкретен, т. е. обеспечивает развитие приспособлений к существующим, а не ко всем возможным условиям среды. Понятно, что развитие способности к полету не очень хорошо совмещается с возможностью быстро бегать. Поэтому птицы, обладающие наилучшими способностями к полету, плохие бегуны. Напротив страусы, которые не способны летать, прекрасно бегают. Приспособление к определенным условиям может быть бесполезно или даже вредно при появлении новых условий. Однако условия обитания меняются постоянно и иногда очень резко. В этих случаях накопленные ранее адаптации могут затруднить формирование новых, что может вести к вымиранию больших групп организмов, как это случилось более 60--70 миллионов лет назад с некогда очень многочисленными и разнообразными динозаврами.
Восторгаясь адаптацией видов к условиям существования, мы часто приходим к неверному заключению о том, что каждая черта строения и функционирования организма имеет существенное адаптивное значение. В действительности дело обстоит не так. Одна из причин этого состоит в целостности организма и плейотропном действии генов. Пусть в данных условиях преимущество получают организмы с ферментной системой, обеспечивающей детоксикацию ядовитых веществ в среде путем их превращения в нерастворимый пигмент. Приспособленность организмов к условиям среды возрастет, однако изменение окраски может не иметь к этому никакого отношения. Хотя в других условиях, как видно в примере с индустриальным меланизмом, окраска бабочек имеет принципиальное значение для выживания вида.
Рассмотрим другой пример: у носорогов Юго-Восточной Азии один рог, а у их африканских родственников два. Нет оснований думать, что это морфологическое различие у носорогов имеет принципиальное значение для адаптации одних в Азии, а других в Африке. Наиболее вероятно, что это "побочный" результат естественного отбора двух исходно различающихся генетических систем. Возникает вопрос: означает ли это, что в последующей эволюции виду легко "расстаться" с признаком, не имеющим прямого адаптивного значения? Как ни странно, сделать это, по-видимому, сложно. Можно прибегнуть к аналогии. Большое ли значение имеет для здания цвет одного кирпича в основании? По-видимому, нет, однако извлечь такой кирпич не просто. Обратимся к эксперименту с дрозофилой -- в ряду поколений был проведен отбор на изменение числа щетинок на брюшной поверхности четвертого и пятого сегментов. В исходной линии число щетинок составляло в среднем 36. Попытки отобрать мух с меньшим числом щетинок были малоуспешными -- такие особи гибли либо оказывались стерильными. Отбор на увеличение числа щетинок оказался более успешным -- за 20 поколений удалось увеличить число щетинок до 56, однако в результате такого отбора стала сильно проявляться стерильность и после прекращения отбора число щетинок за 1--2 поколения стало почти равным исходному -- 39. Один из важных выводов, который следует сделать, состоит в том, что существует немало признаков, не имеющих большого адаптивного значения, их появление в значительной мере случайно, однако они тесно увязаны в, единую систему фенотипа организма.
Суммируя, следует подчеркнуть, что формирование адаптации -- сложный результат взаимодействия факторов эволюционного процесса, где направляющую роль играет естественный отбор, который связует требования среды со структурой генотипа и фенотипа организмов.
20. Искусственный отбор, его особенности и результаты. Современные
методы селекции.