

3. Некротичні реакції і роль системи поліфеноли - поліфенолоксидаза у фітоімунітеті
Некротичні реакції, називані також абортивними і гіперергічними, або частіше надчутливістю, уперше були вивчені більш п'ятдесятьох років тому в явищах стійкості пшениці до іржастих грибів. Показано, що паросткові трубки багатьох іржастих грибів, проникаючи в рослини імунних сортів, досить швидко викликають відмирання клітин рослини-хазяїна, з якими вони вступають у безпосередній контакт. Помічено, що через якийсь час із клітин хазяїна дифундують якісь речовини, які вбивають гіфи гриба. Чим більш високу стійкість має сорт, тим менше зона ураження. У імунних сортів відмирає таке мале число клітин, що проникнення патогену вдається знайти тільки під мікроскопом. У результаті утворюються некротичні плями, розмір яких залежить від тривалості росту гриба і числа убитих клітин рослини-хазяїна (Стекман, Харрер, 1959).
Аналогічна послідовність процесів була встановлена під час ураження різних сортів картоплі, що відрізняються за ступенем стійкості до фітофторозу. Так, відповідно до досліджень Томиіма (Tomiyama, 1955, 1956, 1957), клітина фітофторостійкого сорту відмирає протягом першої години після проникнення в неї інфекційної гіфи паразита. Гіфа ж гриба в клітині гине лише через 10 годин.
Варто відразу ж підкреслити дві обставини.
По-перше, під некрозом у цих реакціях розуміють не узагалі відмирання клітин і тканин, яке відбувається під час розвитку інфекції, а лише «місцеве відмирання клітин під дією токсичних продуктів розкладання» (Гойман, 1954).
По-друге, некроз є лише видимим результатом захисної реакції, і саме його утворення не розкриває причин відмирання рослинних клітин і загибелі паразита, що проникнув у них.
Існує думка, що некротичні реакції захищають рослину тільки від біотрофних паразитів, оскільки такі паразити здатні живитися вмістом тільки живих клітин, і у відмерлих тканинах вони можуть загинути від голоду. З цього погляду для факультативних паразитів, що живляться попередньо убитими клітинами, некротизована ділянка повинна представляти навіть кращий субстрат, чим живі клітини, опір яких необхідно попередньо перебороти.
Експериментальні дані говорять про інше. Показано, що стійкість рослин до ряду факультативних паразитів, і зокрема до В. cinerea, також пов'язана з некротичними реакціями. Загибель паразита за цього настає не через голод, а в результаті отруєння токсичними сполуками, утворюваними у рослині.
Для пізнання захисного механізму некротичних реакцій досить важливого значення набуває вивчення хімічної природи і біологічної дії тих речовин, які виникають у ході цих реакцій. Чи викликають відмирання рослинних клітин і паразита, що проникнув, ті самі речовини, або ж вони є різними?
У літературі з цього питання висловлені різні точки зору. Деякі автори вирішальну роль у некротичних реакціях віддають фітоалексинам, до яких належать речовини різної хімічної природи. Інші розглядають як основний механізм цих реакцій наявну в рослинах систему поліфеноли – поліфенолоксидаза, під дією якої в інфікованій тканині накопичуються продукти окислення поліфенолів, що чинять токсичну дію як на клітини рослини-хазяїна, так і на паразита.
Аказава й Уритани (Akazawa, Uritani, 1961) досліджували зміну ступеня стійкості бульби батата до збудника чорної гнилизни в інтервалі температури навколишнього повітря від 20 до 34° і одночасно простежили за динамікою утворення поліфенолів і фітоалексину – іпомеамарона. Було виявлено, що стійкість батата корелювала з утворенням іпомеамарона, але не залежала від інтенсивності утворення фенольних сполук.
Разом з тим відзначалося, що вміст поліфенолів і активність поліфенолоксидази у відповідь на інфекцію зростає, причому тим сильніше, чим вище стійкість рослин до збудника хвороби. Природно, виникає питання про джерела збільшення вмісту поліфенолів – чи пов'язано воно з новотвором їх або міграцією зі здорових тканин, що примикають. Відповідно до досліджень Озерецковської, Васюкова (1965), сильне збільшення вмісту поліфенолів було виявлено в диску, вирізаному з бульби картоплі, у міру утворення на його поверхні ранової перидерми. Тому що вміст поліфенолів враховувалося у всьому диску, то збільшення їхньої кількості за рахунок перетікання з інших тканин виключалося. Непрямою вказівкою про новотвір поліфенолів у відповідь на поранення служить посилення в заражених і механічно ушкоджених тканинах процесів апотомічного окислення, що являє собою єдиний шлях утворення еритрозо-4-фосфату, необхідного для біосинтезу ароматичних кілець (Рубін, Озерецковська, 1962).
Більшість природних поліфенолів, що зустрічаються в рослинах, має слабку фунгітоксичну дію. Прикладом цьому є фунгітоксичність хлорогенової і кавової кислот, які присутні в здоровій бульбі картоплі. Причому, дія кавової кислоти трохи сильніша, ніж хлорогенової, але в цілому фунгітоксичність їх відносно невелика. Скополетін, аглікон скополіна, також виявленого в тканинах бульби картоплі (Hughes, Swain, 1960), виявився сильним інгібітором росту гіф гриба, але мало пригнічував проростання його спор. Однак тільки на підставі іспиту токсичності фенолів ще не можна судити про захисну роль системи поліфеноли – поліфенолоксидаза. Адже коли мова йде про всю систему, то маються на увазі не тільки самі поліфеноли, але і виникаючі у відповідь на інфекцію продукти їх ферментативного окислення.
Модельними дослідами показано, що продукти окислення поліфенолів мають набагато більш високу фунгітоксичність порівняно з вихідними сполуками. Для їхнього нагромадження повинна бути порушена властива здоровій клітині певна супряженість процесів окислення і відновлення поліфенолів, регульована відповідними ферментами. Ця супряженість процесів і порушується в ході некротичних реакцій. Причиною цьому може бути розрив білково-ліпідної мембрани тонопласту, що оточує клітинну вакуоль, унаслідок чого фенольні речовини виходять у цитоплазму, де піддаються швидкому окисленню (Салькова, Платонова, 1967). Окислені продукти фенолів необоротно пригнічують активність ряду ферментів і в першу чергу дегідрогеназ. Так, у ході некротичних реакцій картоплі було виявлено не тільки посилення активності поліфенолоксидази, але і пригнічення дегідрогеназ, які каталізують відновлення продуктів окислення поліфенолів (Рубін, Аксьонова, 1957). Виявилося, що фунгітоксичність більшості випробуваних сполук фенольної природи помітно зростає після ферментативного окислення. Різна швидкість окислення поліфенолів і різний склад продуктів їх окислення, що утворюються у відповідь на вторгнення паразита у стійких і сприйнятливих сортах, є, очевидно, однією з причин, чому у випадку стійких сортів система поліфеноли – поліфенолоксидаза відіграє захисну роль, а в сприйнятливих сортах не зупиняє розвиток інфекції, хоча і може на якийсь час затримати її просування. Ще Сцент-Дьерди і Кьеторж (Szent-Gyorgyi, Cietorisz, 1931) відзначали, що фунгітоксичність продуктів окислення фенолів укладена в їх неспецифічній дубильній дії на білки, у результаті чого останні стають недоступними для паразита.
Поліфеноли й особливо продукти їх окислення є роз’єднувачами процесів окислювання і фосфорилування. Не виключено тому, що токсична дія цих речовин у некротичних реакціях може бути обумовлено порушенням енергетичного апарата як паразита, так і рослини-хазяїна, унаслідок чого і той і інший позбавляються необхідної енергії для своєї життєдіяльності. Найбільшим роз’єднувачем виявилася кавова кислота, що пригнічувала поглинання неорганічного фосфору наполовину. Хлорогенова кислота, навпроти, майже не впливала на фосфорилування. Продукти окислення обох кислот помітно пригнічували поглинання фосфору. Не виключено, що така дія продуктів ферментативного окислення обумовлюється їх денатуруючим впливом на мітохондриальний білок.
На підставі проведених досліджень можна прийти до висновку, що саме система поліфеноли – поліфенолоксидаза відповідальні у першу чергу за утворення некрозу. Разом з тим, і деякі фітоалексини також можуть роз'єднувати дихання від фосфорилування. Так, іпомеамарон, що міститься у некротизованій тканини батата, ураженого С. fimbriata, порушує енергетику паразита (Uritani, Akazawa, Uritani, 1954). Дуже можливо, що фітоалексини більшою мірою пригнічують процеси обміну паразита та меншою – впливають на рослину. Такої точки зору дотримується Аллен (Alien, 1966), який вважає, що фітоалексини недостатньо токсичні для тканин рослини для того, щоб бути відповідальними за некротичний ефект. Про це ж свідчать результати Круикчанка (Cruickshank, 1963), який знайшов, що в горосі у відповідь на вплив хімічними отрутами фітоалексини утворювались, але виникнення некрозу не спостерігалося.
Таким чином, згідно сучасним уявленням, немає підстави надавати системі поліфеноли – поліфенолоксидаза вирішального значення в захисних реакціях рослини проти паразита, що проникнув, Хоча в ході ферментативного окислення поліфенолів виникають продукти, що значно перевершують за своєю антибіотичною дією вихідні сполуки, їх фунгітоксичність відносно невелика і помітно нижче, ніж в описаних фітоалексинів. Це мимоволі приводить до думки, що в некротичних реакціях поряд із системою поліфеноли – поліфенолоксидаза діє інший механізм, більш активний у відношенні пригнічення самого паразита.
Проведені дослідження ступеня фунгітоксичності екстрактів, витягнутих з некротизованої тканини бульби картоплі (генотип. – R1), що утворилася у відповідь на інфікування несумісною расою фітофтори (раса О), показали, що екстракти, виділені за допомогою спирту, які містять, поліфеноли, помітно уступають за своєю фунгітоксичністю хлороформовим екстрактам, у складі яких поліфеноли відсутні. Ще більш фунгітоксичним виявився ефірний екстракт, що при тих же концентраціях цілком пригнічував проростання спор R. solani.
Хроматографічне розділення ефірних і хлороформових екстрактів методом тонкошарової хроматографії на силікагелі показало, що в тканинах ранового шару, а ще в більшому ступені в некротизованій тканині з'являються нові сполуки, які відсутні у здоровій бульбі.
Останнім часом Томіяма та ін. (Tomiyama et al., 1967) з некротизованої тканини бульби картоплі, яка утворилася у відповідь на інфікування несумісною расою Ph. infestans, виділили й ідентифікували антибіотичну речовину, яка виявилась біоциклічним норсесквитерпеноїдним спиртом приблизно наступної будови:
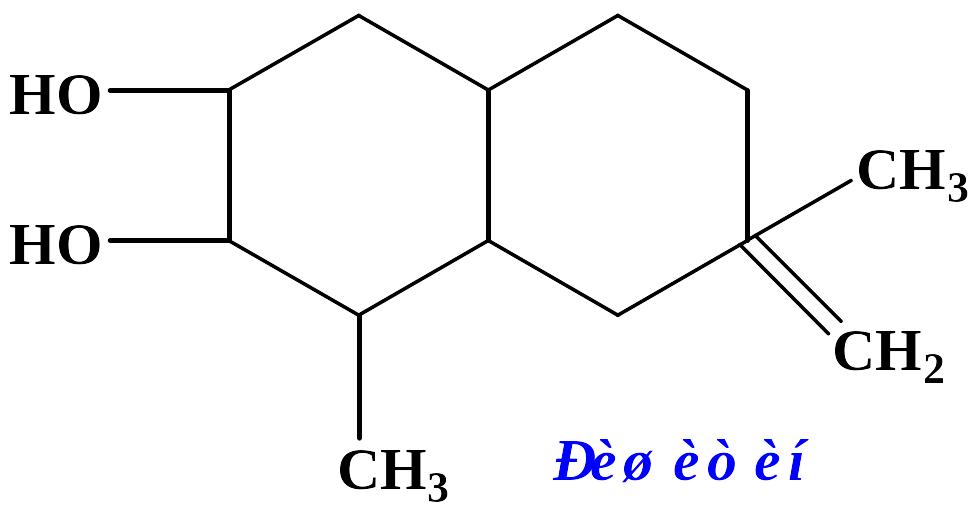
Сполука була названа ришитином. Середня ефективна доза (ЕД50) для неї у відношенні різних рас Ph. Infestans складає 2,7∙10–4М, що відповідає 40 мкг/мл і наближується за силою своєї дії до інших фітоалексинів.
На підставі усього вищевикладеного представляється, що захисна роль некротичних реакцій у стійкості рослин до фітопатогенних мікроорганізмів обумовлена як продуктами окислення поліфенолів, так і іншими антибіотичними речовинами, зокрема фітоалексинам.
Роль продуктів окислення поліфенолів не можна зводити тільки до утворення некрозу. Вони, безсумнівно, діють токсично і на паразита і тим самим можуть послабити його стійкість стосовно фітоалексинів. Крім того, можна чекати синергічної дії тих і інших. Нарешті, продукти окислення поліфенолів можуть одночасно локалізувати вогнище інфекції, захистити його від вторинного зараження, а також попередити проникнення в прилягаючі здорові тканини токсичних речовин, що залишилися на місці інфекції як від загиблого паразита, так і від зруйнованих клітин рослини-хазяїна.
СТІЙКІСТЬ РОСЛИН ДО АНТРОПОГЕННИХ ФАКТОРІВ
ЛЕКЦІЯ № 13
План лекції
Типи атмосферних забруднень.
Компоненти забруднення атмосфери та їх токсичність.
Фізіолого-біохімічні реакції рослин на забруднення повітря.
1. Типи атмосферних забруднень
У країнах з високорозвиненою промисловістю звичайно виділяють два типи атмосферних забруднень, чи «смогу»: лос-анджелеський і лондонський (Томас, 1962). Діючим початком в обох служать газоподібні речовини й аерозолі різного складу.
Лос-анджелеський тип забруднення атмосфери (смог) характеризується вмістом у приземному шарі повітря великої кількості вуглеводнів перекисної природи. Основними джерелами надходження їх в атмосферу служать продукти неповного згоряння бензину й масел у двигунах внутрішнього згоряння і летучі фракції нафти. У районі Лос-Анджелесу, за даними Стенфордського науково-дослідного інституту, у 1953 р. у середньому щодоби викидалося в повітря близько 1080 т вуглеводнів і 1380 т інших органічних забруднювачів. В останні роки кількість викидів зрослася в декілька разів.
Зазначені речовини в момент надходження в атмосферу або в нічний час не чинить дій, які ушкоджують чи подразнюють живі організми. Їхня токсичність різко зростає після деякого періоду освітлення сонячними променями. Відповідно до теорії Хаагена-Смита, утворення смогу відбувається в результаті фотохімічної взаємодії його компонентів з атомарним киснем. Двоокис азоту, сірчистий ангідрид і альдегіди здатні поглинати ультрафіолетові промені і переходити в активний стан. Вивільнюваний при цьому атомарний кисень реагує з молекулярним киснем повітря. Частина атомарного кисню може утворюватися із сірчистого ангідриду чи альдегідів. В утворенні озону найбільш істотна роль належить двоокису азоту. Молекула останнього під дією ультрафіолетових променів перетворюється в окис азоту й атомарний кисень. Передбачається, що реакції здійснюються в такий послідовності:
NO2 + hν = NO + O,
O + O2 = O3,
NO + O2 = NO3,
NO3 + O2 = NO2 + O3.
Цей процес повторюється багаторазово за збереження кількості окислів азоту, що беруть участь у ньому. Тому навіть незначний вміст окислів азоту в повітрі служить причиною нагромадження в ньому озону, що досягає концентрації 3-4∙10-5% і більше від об’єму повітря.
Озон і окисли азоту реагують з органічними домішками повітря, у результаті чого утворяться ненасичені вуглеводні, альдегіди, кетони, кислоти й інші речовини із сильно вираженою фітотоксичною дією. Вважають, що пероксилацетилнітрат (ПАН), який утворюється за цих умов, є специфічним діючим початком смогу лос-анджелеського типу.
За утворення надлишку озону в повітрі відзначене подальше окислення двоокисів азоту і сірки з утворенням відповідно азотної і сірчистої кислот:
2NO2 + O3 = N2O5 + O2,
N2O5 + H2O = 2HNO3.
Кислоти, що утворилися, утримуються в повітрі у виді рідкого аерозолю. Контактуючи з листям, вони викликають точкові ушкодження.
Хімічний склад лос-анджелеського смогу характеризується великою кількістю насичених і ненасичених вуглеводнів рядів від C2 до C10, що містяться в ньому.
Лос-анджелеський смог широко розповсюджений у багатьох містах США, Італії, Франції й інших країн з інтенсивним автомобільним рухом, але ослабленим обміном повітря і достатком сонячного світла.
Лондонський тип забруднення атмосфери (смог) характеризується скупченням у приземному шарі повітря переважно продуктів неповного згоряння кам'яного вугілля.
Сірчистий і сірчаний ангідриди легко розчиняються у крапельках тумана й утворюють аерозолі сірчистої і сірчаної кислот. Пилоподібні частки вугілля, золи і різних солей також мають фітотоксичну дію. Найчастіше речовини, що містяться в лондонському смозі, мають відновні властивості. Висока токсичність останнього виявляється під час штилю і температурної інверсії, але наявність тумана не обов'язкова.
Атмосферне забруднення лондонського типу властиво не тільки Великобританії, але і промисловим районам інших країн північної Європи, при викиданні атмосферу речовин ідентичного складу і за подібних умов – високої вологості повітря, часто повторюваної суцільної хмарності, температурній інверсії і слабкому обміні повітря в приземному шарі.
Промисловість України – високорозвинена і багатогалузева. Особливо інтенсивно в країні ведеться видобуток і переробка кам'яного вугілля, залізної й інших руд, нафти, природного газу. В Україні працює велика кількість різноманітних хімічних підприємств і заводів по переробці сільськогосподарської продукції. Так, на одиницю площі в Україні в 1961 р. продуктивність підприємств чорної металургії перевершувала в 2-4 рази ті ж показники в США. В наступні роки це розходження ще більш зросло.
Кислі і лужні гази, реагуючи між собою, а також з активними компонентами повітря (водяна пара, кисень і ін.), утворюють нові сполуки. Усі вони в газоподібному чи розчиненому у воді виді проникають у рослини, викликаючи порушення їхньої життєдіяльності. Іноді шкідлива дія токсичних газів і аерозолів виявляється на відстані до 50 км від підприємств.
На Україні порівняно рідко сполучається безвітряна погода з тривалою температурною інверсією і суцільними хмарами, туманами чи опадами. У зв'язку з цим протягом вегетаційного періоду тут рідше, ніж у північних районах, спостерігається скупчення основної маси фітотоксикантів у приземному шарі повітря. Цьому протидіє порівняно рівнинний рельєф місцевості, а також постійні вітри. Якщо ж сполучається затишність з туманом чи опадами, то у земної поверхні утримується утворена промисловими і транспортними викидами імла з різким дратівним запахом, що спричинює сильну токсичну дію на рослини.