Тести з теми “Основи молекулярної генетики ”
1.Із нітратів , нітритів і нітрозамінів в організмі утворюється азотиста кислота , яка зумовлює окисне дезамінування азотових основ нуклеотидів . Це може призвести до точкової мутації - заміни цитозину на:
A.Гіпоксантин
B.Гуанін
C.Тимін
D.Аденін
E.Урацил
2. У хворих на пігментну ксеродерму шкіра надзвичайно чутлива до сонячного світла , може розвиватись рак шкіри . Причиною є спадкова недостатність ферменту УФендонуклеази . Внаслідок чого цей порушується процес ?
A.Репарації ДНК
B.Зворотньої транскрипції
C.Реплікації ДНК
D.Транскрипції
E.Трансляції
3. Для лікування урогенітальних інфекцій використовують хінолони - інгібітори ферменту ДНК-гірази . Укажіть , який процес порушується під дією хінолонів у першу чергу.
A.Репарація
B.Реплікація
C.Ампліфікація генів
D.Рекомбінація генів
E.Зворотна транскрипція
E.Трансляції
4.На судово -медичну експертизу надійшла кров дитини та передбачуваного батька для встановлення батьківства . Вкажіть , ідентифікацію яких хімічних компонентів необхідно здійснити в дослідній крові.
A. Гя-РНК.
B.т-РНК.
C.р-РНК.
D.ДНК.
E.мя-РНК.
5. Молекулярний аналіз гемоглобіну пацієнта , що страждає на анемію , виявив заміну 6Глу на 6Вал бета-ланцюга . Який молекулярний механізм патології ?
A.Генна мутація
B.Хромосомна мутація
C.Геномна мутація
D.Ампліфікація генів
E. Трансдукція генів |
Еталони відповідей : |
152 |
1- Е; 2 – А; 3 – ВД; 4 – Д;5 – А. |

Механізми взаємодії гормонів з клітиною
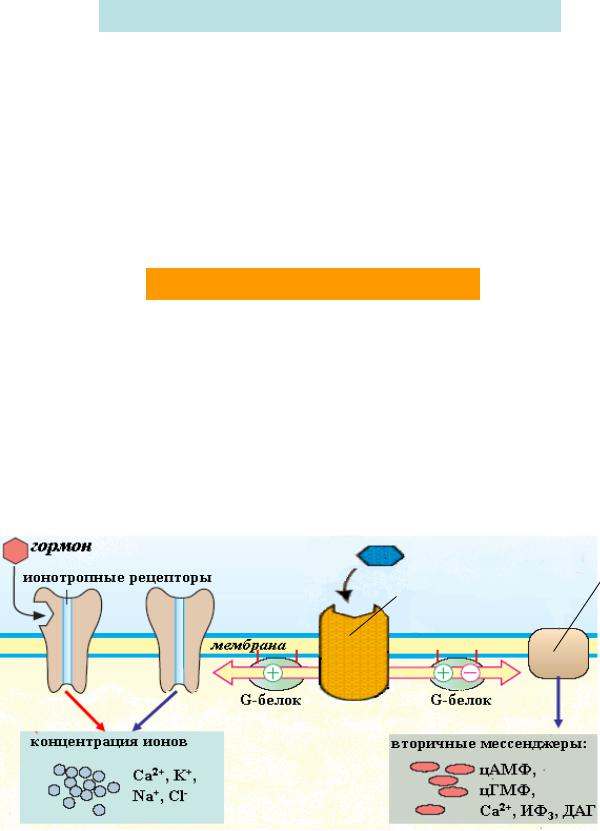
Усі гормони поділяють на дві великі групи в залежності від клітинної локалізації рецептора (на клітинній мембрані або в клітинні ), з яким взаємодіє гормон . Це, в свою чергу, залежить від структури та властивостей самого гормону :
гідрофільні гормони – білково -пептидної природи та похідні амінокислот – не здатні проникати всередину клітини -мішені крізь ліпофільну клітинну мембрану , діють на клітину за рахунок зв'язування з мембранним рецептором ;
ліпофільні гормони – стероїдні гормони і йодтироніни – проникають в клітину - мішень і взаємодіють із специфічними рецепторами в цитоплазмі або в клітинному ядрі.
Механізм дії гідрофільных гормонов
Трансформація гормонального сигналу в специфічну відповідь клітини -мішені
відбувається в результаті таких молекулярних подій:
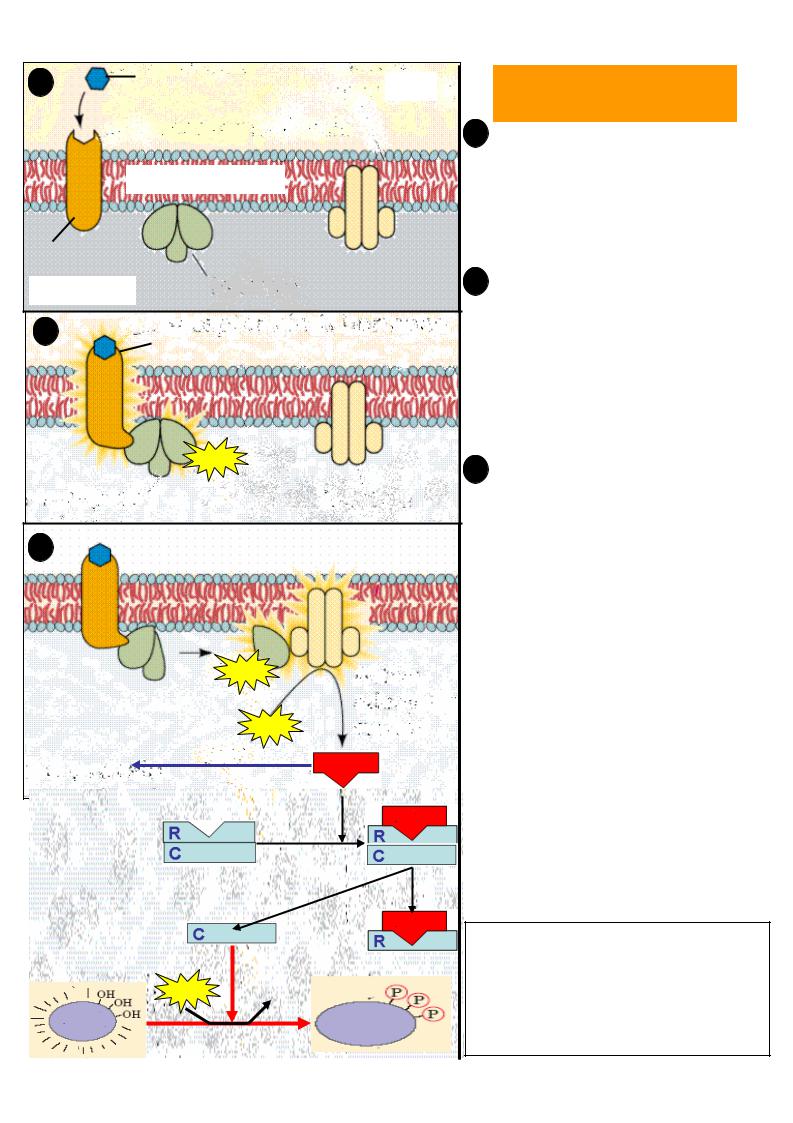
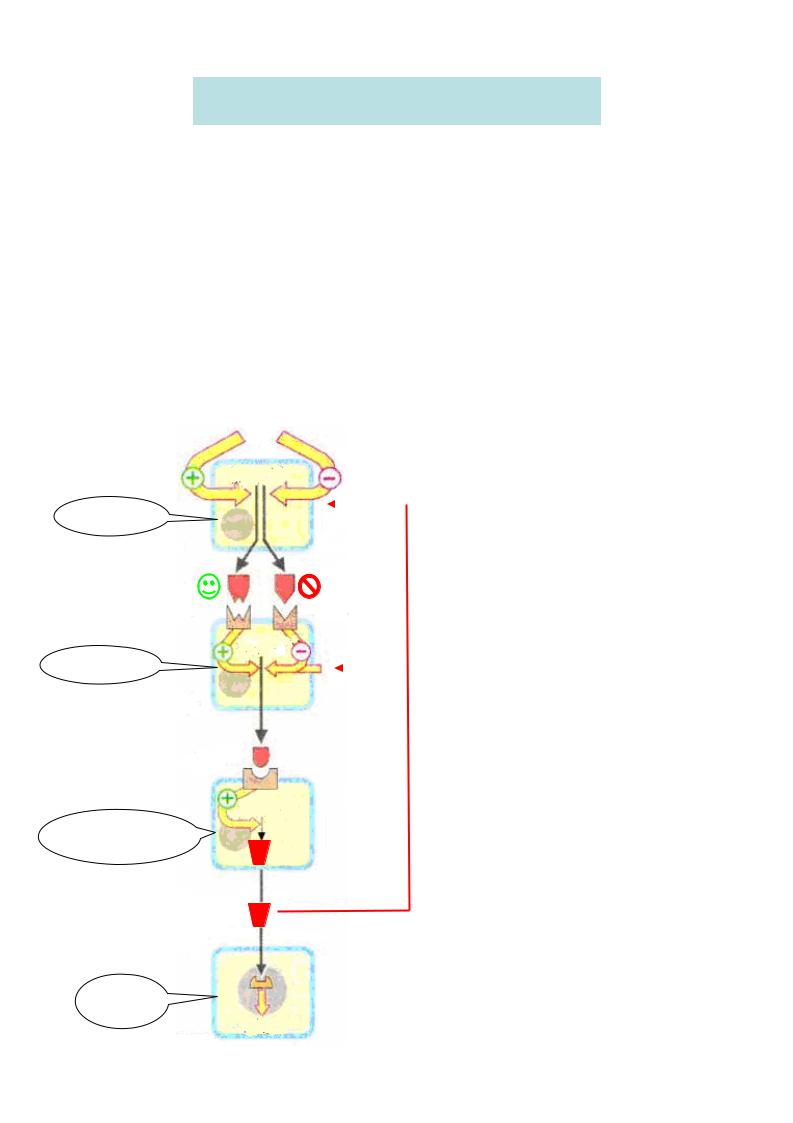
Передача гормонального сигналу здійснюється за допомогою білкових рецепторів , розташованих на поверхні плазматичної мембрани . Розрізняють два основних типи мембранних рецепторів : іонотропні рецептори – після взаємодії з гормоном вони сприяють відкриттю іонних каналів на мембрані та розвитку швидких іонних потоків (кальцію , натрію , калію, хлору);
метаботропні рецептори активують системи клітини за допомогою |
білків- |
||
трансдукторів (G-білків). |
|
|
|
гормон |
ефекторні |
|
|
метаботропний |
ферменти : |
||
аденілатциклаза , |
|||
рецептор |
|||
гуанілатциклаза , |
|||
|
|||
|
фосфоліпаза С, |
||
|
фосфоліпаза А2. |
||
G-білки - це внутрішньомембрані білки, які приймають хімічний сигнал від рецептора , модифікованого гормоном , та передають цей сигнал на ефекторні системи клітини . Існує декілька типів G-білків, основні з них:
Gs – стимулюють аденілатциклазу ; |
|
Gi - інгибують аденілатциклазу ; |
|
Gq – активують фосфоліпазу C. |
157 |

Ефекторні ферменти – аденілатциклаза , гуанілатциклаза , фосфоліпаза Ста ін.,
локалізовані на внутрішній поверхні клітинної мембрани . Вони стимулюють утворення в цитоплазмі клітини вторинних посередників – месенджерів.
Найбільш важливими вторинними месенжерами є: циклічний АМФ (цАМФ )
2+ , інозитолтрифосфат ІФ 3, діацилгліцерид
Подальша передача гормонального сигналу від вторинних месенжерів здійснюється за допомогою протеїнкіназ – ферментів , які фосфорилюють білки (більше 100 різних білків, у тому числі ряд ферментів і факторів транскрипції ). У результаті фосфорилювання змінюється функціональна активність цих білків.
|
|
|
|
|
|
|
|
|
|
|
|
|
Ліпофільні гормони |
||
|
|
|
|
|
|
|
|
|
|
|
|
|
|||
|
|
|
|
|
Гідрофільні гормони |
||||||||||
|
|
|
|
|
діють за допомогою |
||||||||||
|
|
|
|
|
діють за допомогою |
||||||||||
|
|
|
|
|
внутрішньоклітинних |
||||||||||
|
|
|
|
мембранних рецепторів |
|||||||||||
|
|
|
|
рецепторів |
|||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|||
|
|
|
|
|
|
|
|
|
|
|
|
|
|||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Гормон |
|
Гормон |
Гормон |
|||||||||
|
|
(білково -пептидні гормони , |
(інсулін , СТГ) |
(стероїдні гормони , |
|||||||||||
|
|
|
адреналін) |
Ауто- |
|
|
|
йодтироніни ) |
|||||||
|
|
|
|
||||||||||||
|
|
|
|
|
|
|
|
|
|
|
Рецептор ( |
|
усередині клітини ) |
||
|
|
|
|
|
|
|
|
|
|
|
|
||||
|
|
|
|
|
|
|
|
|
|
||||||
|
|
|
|
|
|
|
|
фосфорил |
|
||||||
|
|
Рецептор (у мембрані ) |
|||||||||||||
|
|
ювання |
|
|
|
||||||||||
|
|
|
|
|
|
|
|
|
|
|
|||||
|
|
|
|
|
|
|
|
рецептора |
Комплекс «гормон – рецептор » |
||||||
|
|
|
G – білок |
|
|
|
|
|
|
|
|||||
|
|
|
|
|
|
|
|
|
|
||||||
|
|
|
|
|
|
|
|
|
|
|
|
||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Ефекторний |
|
фермент |
|
G – білок |
|
Транспорт |
|
комплексу |
|||||
|
|
|
|||||||||||||
|
|
(аденілатциклаза , |
|
|
|
|
|
«гормон - рецептор » у ядро |
|||||||
|
|
фосфоліпаза С, та ін.) |
|
|
|
|
|
|
|
|
|||||
|
|
|
|
|
|
|
|
|
|||||||
|
|
|
|
|
|
|
|
|
Каскад |
Взаємодія |
|
комплексу |
|||
|
|
|
|
|
|
|
|
|
|
||||||
|
|
|
|
|
|
|
|
|
|||||||
|
|
|
|
|
|
|
|
фосфорилювання |
«гормон - рецептор » с ДНК |
||||||
|
|
Вторинний месенжер |
|
||||||||||||
|
|
|
|
білків |
|
|
|
||||||||
|
|
(цАМФ , Са 2+, ІФ , ДАГ) |
|
|
|
|
|
|
|
|
|
||||
|
|
|
|
|
|
|
|
|
|
|
|||||
|
|
3 |
|
|
|
|
|
|
|
|
|
||||
|
|
|
|
|
|
|
|
|
|
|
|
|
Індукція або репресія |
||
|
|
|
|
|
|
|
|
|
|
|
|
|
|||
|
|
|
|
|
|
|
|
|
|
|
|
|
транскрипції |
||
|
|
|
|
|
|
|
|
|
Активація |
||||||
|
|
Протеїнкінази |
|
||||||||||||
|
|
|
|
|
|
||||||||||
|
|
|
|
||||||||||||
|
|
|
|
|
|
|
|
ферментів та |
|
|
|
||||
|
|
|
|
|
|
|
|
|
|
|
|||||
|
|
|
|
|
|
|
|
|
факторів |
|
|
|
|||
|
|
|
|
|
|
|
|
Трансляція - |
|||||||
|
|
Фосфорилювання білків |
транскрипції |
синтез білків (ферментів ) |
|||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Зміна функціональної активності білків |
Зміна кількості |
158 |
(ферментів ) |
білків (ферментів ) |
Поява в клітині вторинного посередника є пусковим моментом для зміни метаболізму , який здійснюється , як правило , шляхом фосфорилювання білків. Роль вторинних посередників можуть виконувати цАМФ , цГМФ , інозитолтрифосфат , діацилгліцерин Ca2+. Найпоширенішим і добре вивченим вторинним посередником є циклічний 3',5'-
аденозинмонофосфат (цАМФ ):
аденілат - циклаза
фосфодіестераза
АТФ |
цАМФ |
АМФ |
Зв'язування гормону з рецептором активує аденілатциклазу і, отже, призводить до підвищення внутріклітинної концентрації цАМФ, що веде до збільшення швидкості фосфорилювання білків. Наявність каскаду ферментативних реакцій між зв'язуванням гормону з рецептором і зміною метаболізму дозволяє значно підсилити первинну дію гормону . Таким чином , відбувається багатократне посилення гормонального сигналу – ампліфікація: на одну молекулу гормону припадає до 100 молекул цАМФ ). Припинення дії гормону в даному випадку досягається руйнуванням вторинного месенжера – за допомогоюфосфодіестерази він перетворюється на АМФ.
Інший ефекторний фермент , що знаходиться в ліпофільній мембрані фосфоліпаза– С– гідролізує ті фосфоліпіди (фосфоінозітолдіфосфати ,) що знаходяться в цій самій мембрані , внаслідок чого утворюються інші вторинні внутрішньоклітинні посередники : гідрофільнийінозитолтрифосфат і ліпофільний діацилгліцерид .
діацилгліцерид (ДАГ)
|
|
фосфоліпаза С |
|
159 |
фосфоінозитолдіфосфат |
(ФІФ) |
інозитолтрифосфат (ІФ) |
3 |
|||
|
2 |
|
|