

МОЛЕКУЛЯРНІ АСПЕКТИ ПЕРЕДАЧІ ГЕНЕТИЧНОЇ ІНФОРМАЦІЇ
" Генетична і нформація зберігається " в клітинних ядрах. Вона " зашифрована у вигляді послідовності"нуклеотидів"в"первинній"структурі"ДНК."
Первиння(структура(ДНК(
– це певна послідовність дезоксирибонуклеотидів в полінуклеотидному ланцюзі , зв'язаних 3’,5’- фосфодиефірними зв'язками .
На 5’-кінці знаходиться фосфатна група
на 3’-конце – ОН-група пентози .
Вторина(структура(ДНК!
На 5’-кінці знаходиться фосфатна група на 3’-кінці – ОН-група пентози .
- це подвійна спіраль , утворена двома полінуклеотидними ланцюгами , закрученими навкруги загальної осі і розташованими антипаралельно. Ланцюги утримуються за рахунок водневих зв'язків між нуклеотидами ,
що комплементарні -А -Т і G-C, та
лежать в одній площині (перпендикулярної осі). Спіраль стабілізована гідрофобними взаємодіями між основами .
Ланцюги комплиментарні , але не ідентичні один одному , їх склад розрізнений.
131
• |
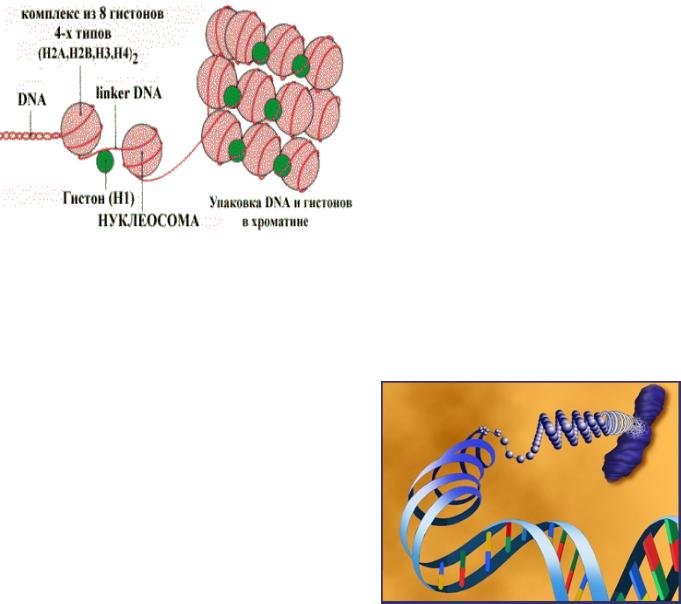
Кожна молекула ДНК упакована в окрему хромосому , яка містить |
|
також різноманітні білки, тобто ДНК знаходиться в клітині в комплексі з |
• |
білками , утворюючи нуклеопротеїн - хроматин або хромосоми . |
Отже, в кожній хромосомі знаходиться одна гиганська ДНК, в |
|
|
середньому що містить 150 млн нуклеотидних пар, довжина молекули |
|
3-5 см. Величина хромасомы - 1нм, тобто ДНК суперспіралізована , |
|
лінійний розмір кожної молекули ДНК зменшується в 8000 разів! |
Склад(хроматину:(
! ДНК"="30=40%" "гістоны(="30=50%""="основні"білки""""
невеликого"розміру"з"високим"вмістом"
позитивно"заряджених"амінокислот"="
аргініну"і"лізину."Є"5"типів"гистонів:"" Н2А,"Н2В,"Н3,"Н4"="утворюють"нуклеосоми,( які"складаються"з"146"пар"нуклеотидів" ДНК"(заряджених"негативно)"і"восьми" гістонів,"що"мають"позитивний"заряд."Н1"="" зв'язуються"з"ДНК"в"міжнуклеосомних" ділянках"(лінкерних"послідовностях)"і" захищають"їх"від"дії"нуклеаз "
" негістонові( білки" (кислі)=" " 4=33%" " =це" регуляторні"білки"та"ферменти " РНК"="1.5=10%"
" Гаплоїдний" геном" кожної"
клітини" людини" включає" 3,5" 10" " пар" основ" (23" пари" хромосом)" достатньо" для" 1,5" млн" пар" генів," а" кодується" тільки"100000"білків"
132
Функції(ДНК"
Генетична"інформація,"що"закладена"в"ДНК,"служить"двом"метам:! 1!="передача"генетичної"інформації"дочірньої"ДНК"при"розподілі"клітини,"тобто"
передача"самої"себе"у"ряді"клітинних"поколінь"і"поколінь"організмів;! 2!="для"біосинтезу"білкових"молекул,"тобто"передача"генетичної"інформації"на" РНК"в"процесі"життєдіяльності" Обидві"функції"засновано"на"тому,"що"молекула"ДНК"служить"матрицею:"в" першому"випадку"для"реплікації,"в"другому"="для"транскрипції.""
Нуклеїнові кислоти і білки називають інформаційними молекулами , оскільки в чергуванні їх мономерів закладено певне значення . Послідовність нуклеотидів в ДНК визначає структуру всіх білків клітки. Ділянки ДНК (гени), кодуючі певні білки, копіюються (транскрибіруються ) у вигляді полінуклеотидного ланцюга матричної РНК, яка потім служить матрицею для синтезу білка. Таким чином , генетична інформація , записана в ДНК (в генотипі ) забезпечує утворення фенотипічних ознак клітини , тобто генотип трансформується у фенотип . Цей напрямок потоку інформації включає три типи матричних синтезів .
Потік(генетичної(інформації:""""""""""""""""""""""""""""""""""""""""
1" |
|
""рРНК |
|
|
|
2" |
|
3" |
білок!! |
||
!!!ДНК |
! мРНК! |
||||
|
|
"""""""""""""""""""""""""""""""""""" тРНК"
1!–!репликація="""синтез"ДНК"на"матриці"ДНК"
2!:!транскрипція=синтез"РНК"на"матриці ДНК,"
зворотня(транскрипція"="синтез"ДНК"на""""""""
""""""матриці"РНК"(онковіруси)"
3!:!трансляція!!:!!синтез"білка"на"матриці"РНК"""
Експресія!генів:!
транскрипц ія" трансляц ія"
ДНК!
РНК!
білок!
133
РЕПЛІКАЦІЯ – процес подвоєння ДНК, суть якого складається в утворенні ідентичних копій ДНК для передачі генетичної
інформації в поколіннях клітин та організмів .
Принципы репликации :
Комплементарність Антипаралельність Уніполярність Потреба в затравці Преривистість
Полуконсервативність
1"
2
3
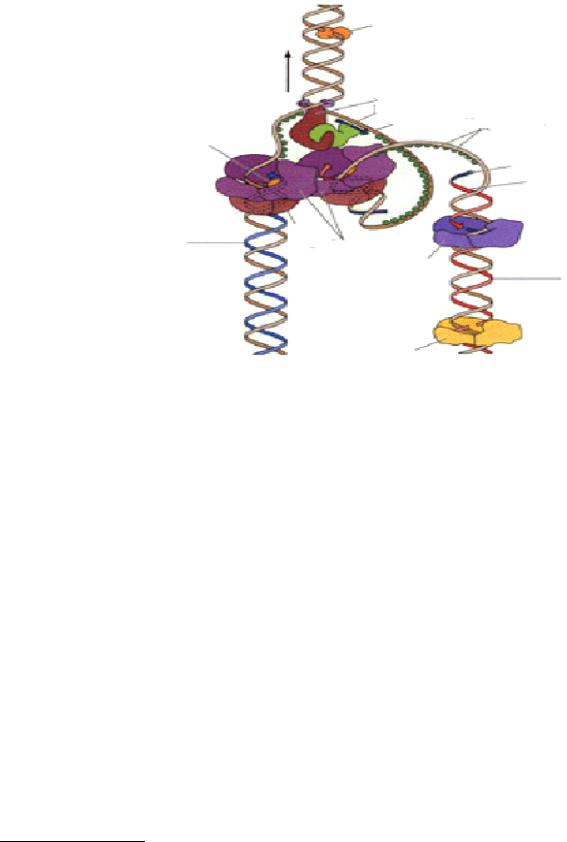
Реплісома-" |
( мультиферментний" |
5" |
4" |
комплекс" |
=" ДНК=репліказная" |
|
|
|
|
система, яка " включає біля " 20" ферментв"і"білкових"факторів:"
6
1 - топоізомераза (гіраза) - зворотня нуклеаза , спочатку вона розриває ланцюг ДНК, а після закінчення реплікації зашиває тимчасові надрізи , розкручує суперспіраль ДНК і виконує “шарнірну функцію ” - перешкоджає обертанню суперспіралізованого ланцюга , (вносить одноланцюгові розриви в ланцюг ДНК і тут же її репарує .)
2- хеліказа - використовує енергію АТФ для розплітання подвійної спіралі ДНК, розриває водневі зв'язки між компліментарними нуклеотидами в
подвійному ланцюзі ДНК (застібка “блискавка ”)
3 - ДНК-залежна РНК-полімераза (праймаза ) синтезує- праймер -
олігорибонуклеотид , який є затравкою для ДНК-полімерази III
4 - ДНКполимераза I (фермент Корнберга)- розщеплює праймер (володіє нуклеазною активністю ) і на його місці синтезує компліментарний ланцюг ДНК (володіє полімеразною активністю ), заповнює проломи .
5 - ДНК-полімераза III -основний фермент реплікаці-ї синтезує компліментарні дочірні ланцюги на материнських ланцюгах , що
розійшлися , у напрямі 5’ → 3’. Вона не здатна ініціювати синтез нового ланцюга ДНК, може лише подовжувати затравку -праймер . Утворює 3,5- фосфоефірні зв'язки між нуклеотидами , використовуючи як субстрати нуклеозидтрифосфати (НТФ).
6 - ДНК-лігаза зшиває фрагменти синтезованої дочірньої ДНК.
134
Сумарне!рівняння!реплікації:!
""""""n"дАТФ""""""""""""""ДНК=матрица"""""""""""""""" "дАМФn"
""""""n"дГТФ" |
ДНК |
дГМФn" +"(n+n+n+n)"ФФ" |
""""""n"дЦТФ""""""""""ДНК=полімераза"""""""""" |
"""""дЦМФn""""""""""""""" |
|
""""""n"дТТФ""""""""""""""""""""""""""""""""""""""""""""""""""""""""дТМФn"
Синтез( кожного( дочірнього( ланцюга( ДНК( іде комплементарно( та антипараллельно до(матричного(ланцюга , та завждив(напрямку 5'((((((((→3'.((
Для"реплікації є необхідним" ряд"умов:"
наявність" дезоксирибо= нуклеотидтрифосфатів" (дАТФ," дГТФ," дЦТФ." дТТФ)," як" структурного" матеріалу" при" збірці"нових"ланцюгів"ДНК"і"як джерела"енергії"
розплетення подвійної " спіралі" ДНК," формування реплікативної"вилки"
утворення " “затравки”" –" праймера"
наявність ферментів" |
та" |
|
специфічних" білків, |
|
що |
приймають участь |
" |
в" |
реплікації"(реплісоми)" |
|
|
на явність" ионів" магнію, що є необхідними для" дії" всіх" полімераз"
135
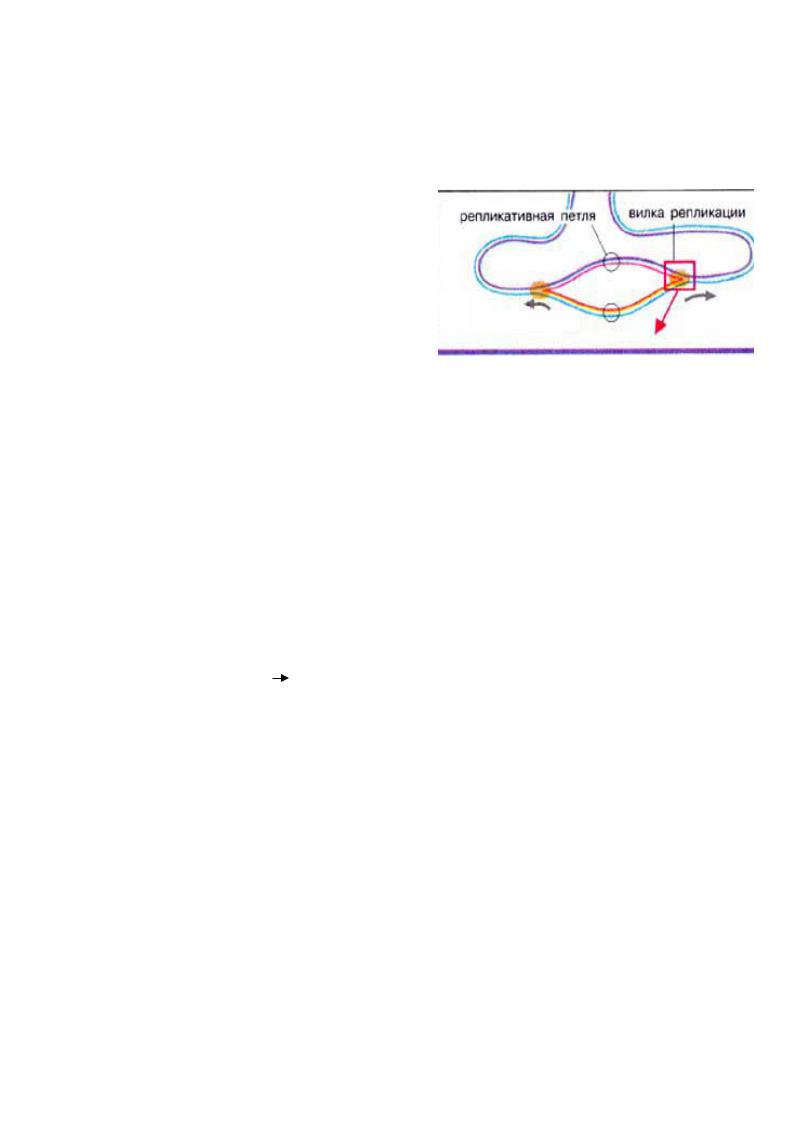
Наявність дезоксирибонуклеотидтрифосфатів (дАТФ, дГТФ, дЦТФ , дТТФ), як структурний матеріал для нових ланцюгів ДНК і як джерела енергії . В даний час процес реплікації у прокариот достатньо вивчений , тоді як багато аспектів реплікації еукаріот залишаються неясними . Проте з великою часткою вірогідності можна стверджувати , що в більшості клітин цей процес протікає подібно
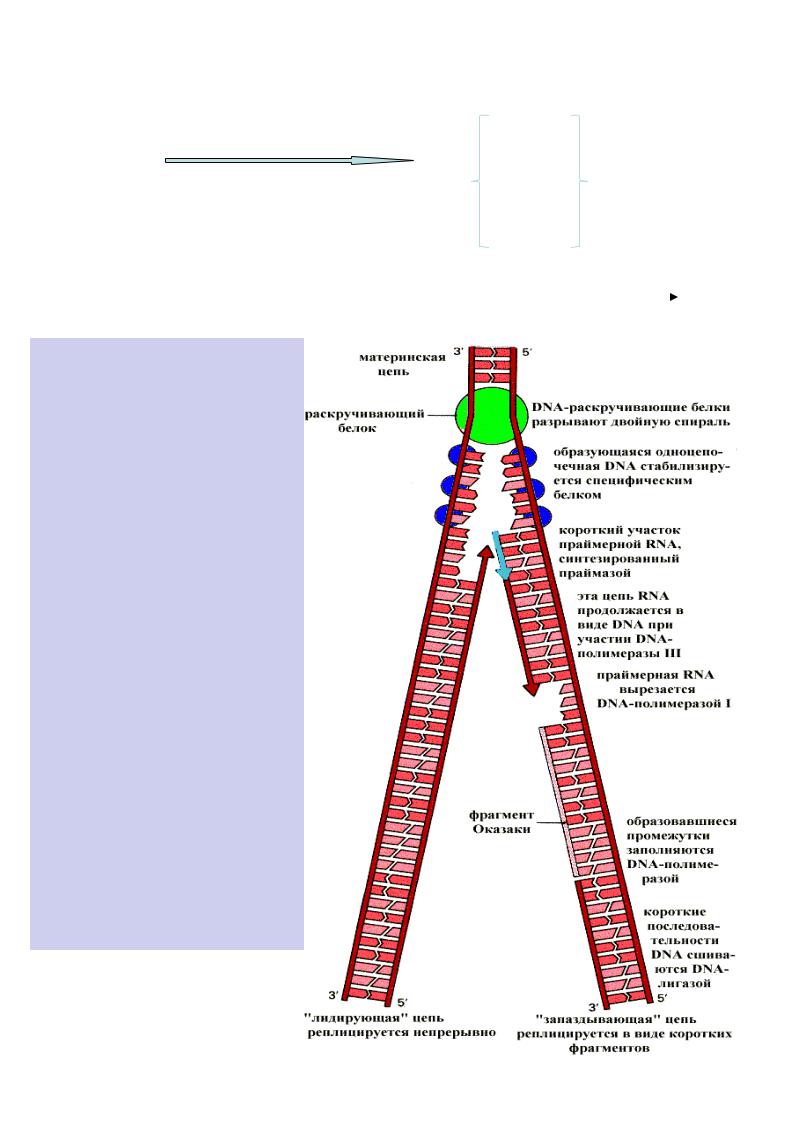
В бактеріях реплікація починається із специфічної крапки в кільцевій ДНК (область початку реплікації ) і продовжується в обох напрямах . В результаті утворюються дві репликативні вилки, які просуваються в протилежних напрямках , тобто обидва ланцюги репліцируються одночасно
1.!Ініціація!!! "ЭТАПИ(РЕПЛІКАЦІЇ(!"
ДНК-полімераза не здатна починати синтез нового ланцюга з її першого нуклеотида . Вона може подовжувати вже наявний ланцюг , тому для початку реакції потрібнаРНКприманка (праймер ), якою є короткий олігорибонуклеотид (100-200 нуклеотидов ), комплементарний до матричного ланцюга ДНК. Синтез починається з реакції між 3’- ОН групи РНК-приманки та 5’-ОН-фосфатної групи дезоксинуклеозидтрифосфата з видаленням пірофосфату . Вибір чергового нуклеотида на кожному кроці синтезу
визначається матричним ланцюгом ДНК згідно правилу комплементарности (Уотсон і Крик, 1953 р.)"
2.!Елонгація!
Реплікація на двох антипаралельних ланцюгах відбувається одночасно . Проте , у
напрямку 3’→5’ синтез йти |
не може, тому на одному ланцюзі - |
лідируючому - |
|
полімеризація йде безперервно , а на другому – |
що запізнюється |
- синтез йде |
|
фрагментами Оказаки по 150 - 200 нуклеотидів проти руху реплікативної вилки. Кожний такий фрагмент містить праймер , який видаляє ДНК-полімераза I, поступово відрізуючи від 5’-кінця поодинці рибонуклеотиди , а до 3’-концу фрагмента вона приєднує дезоксирибонуклеотиди , заповнюючи пролом . Зшиває фрагменти ДНКлігаза."
3.!Термінація""
Закінчується реплікація тоді, коли вся матриця скопійована . В утвореній таким чином подвійній спіралі ДНК тільки один з ланцюгів синтезований наново . Тому говорять , що
реплікація ДНК відбувається понапівконсервативному механізму. У E.coli петля росте із швидкістю 1000 пара нуклеотидів за секунду . Для реплікації всієї молекули хромосомної ДНК цієї бактерії необхідні 42 хвилини . У еукаріот реплікативна петля
росте із швидкістю 100 п.н. в секунду . Ймовірно , це пов'язано з наявністю зв'язку з гістонами . У людини для повної дуплікації ДНК необхідні 8 годин."
136

РЕПАРАЦІЯ ПОВШКОДЖЕНЬ ТА ПОМИЛОК ДНК
Передача"генетичної"інформації"в"неспотвореному"вигляді"="найважливіша"умова"
збереження"вигляду."Це"забезпечується"за"рахунок"комплементарности."Проте,"
можливі"як"помилки"реплікації,"так"і"несприятлива"дія"навколишнього"середовища" (радіація,"УФ=опромінення,"хімічні"агенти).""
Існує"спеціальна"система"моніторингу"точності"спаровування"нуклеотидів,"причому"
здійснюється"подвійна"перевірка:""
="при"включенні"ДРН"в"ланцюг,"що"росте"
="після"включення"ДРН"шляхом"видалення"помилкових"нуклеотидів"(помилки" відбуваються"не"частіше,"ніж"1"разів"на"10""пар"основ).
((((((((((((((((((((((((Причини(спонтанних(порушень
" помилки"репликації
"депурин ізація"з-за слабкої"N=гликозидного"зв'язку в"пуринах" "дезам інування "(Ц У,"А Г,"Г Х)"
"Причини(індукованих(порушень(
"алк ілірування"азотових"основ"(алкілюючі агенти"застосовуються як інгібітори" новоутворувань)"
утворення "зшивок"="піримідинових"димеров"між"сусідніми"основами"(тиміном)"під" дією УФ=опромінення"
Репарація!–!видалення"пошкоджених" ділянок"ДНК"або"помилково"вбудованих" нуклеотидів"в"результаті"дії"спеціальних" ендо"="та"екзонуклеаз.""
Этапи(репарації:"
1) впізнавання"місця"пошкодження" розщеплювання"зв'язку"ендонуклеазою""
2)"добудова"="“латка”"=""пошкодженого"ланцюгу"
ДНК=полімеразою"
3)"відщеплення"ушкодженої"ділянки"
ендонуклеазою""
4)"зшивання"3=кінця"латки"з"5=кінцем""основного"
ланцюга"ДНК=лігазю
137
Порушення репарації УФ-індукованих пошкоджень ДНК (порушення синтезу УФ-специфічної ендонуклеази ) приводить до сублетального спадкового захворювання –пігментної ксеродерми.
Генні мутації
Зміни генетичної програми ДНК клітин - це мутації . Розрізняють хромосомні мутації (зміна числа хромосом , хромосомні аберації ) і молекулярні або генні мутації .
Генні мутації - це успадковані зміни первинної структури ДНК, які ведуть або до припинення синтезу білка, або до синтезу зміненого , дефектного білка.
Мутації в регуляторних ділянках оперона ведуть до порушення регуляції або до припинення синтезу ДНК.
Існують наступні варіанти генних мутацій: Транзиція - заміна пар основ
Місенс -мутація - приводить до зміни значення кодону при заміні нуклеотида , а, значить , до синтезу зміненого білка. Наприклад , серповидно -клітинна анемія : кодон, що відповідає за включення глу в ланцюг гемоглобіну , перетворюється на кодон вал.
Заміна одного нуклеотида не завжди веде до зміни значення кодону (оскільки код вироджений ) - така зміна ДНК фенотипічно не виявляється.
Нонсенс -мутація - в результаті заміни образуется один з термінуючих кодонів , при цьому синтез белка препиняється та утворюється незавершений білок.
Делеція - випадіння однієї пари або групи основ
Вставка однієї пари або групи основ. Ці мутації можуть бути як із зміною рамки зчитування (випадання або вставка одного або двох нуклеотидів ), що веде до синтезу безглуздого білка, так і без зміни рамки зчитування (три нуклеотиди ) - синтезується білок, вкорочений на 1 амінокислоту.
Зміна місцеположення окремих ділянок ДНК.
138
Транскрипція – це синтез всіх видів РНК по матриці ДНК, який здійснюється ферментом ДНК-залежною РНК-полімеразою .
Молекула ДНК, що береже генетичну інформацію , безпосередньої участі в синтезі білка не приймає , але інформація , що записана в послідовності нуклеотидів ДНК, копіюється (транскрибується ) у вигляді м-РНК з подальшою трансляцією в поліпептидний ланцюг білка. Транскрипція , як і реплікація ДНК, - ендергічний процес , зв'язаний з використанням нуклеозидтрифосфатів (АТФ, ГТФ, УТФ, ЦТФ) в якостісубстратів та джерел енергії.""
Принципи(транскрипції:((
""" Комплементарність."" """А нтипаралельність.!" """ Уніполярність.!"
""" Беззатравочність."" """ Асиметричність.!"
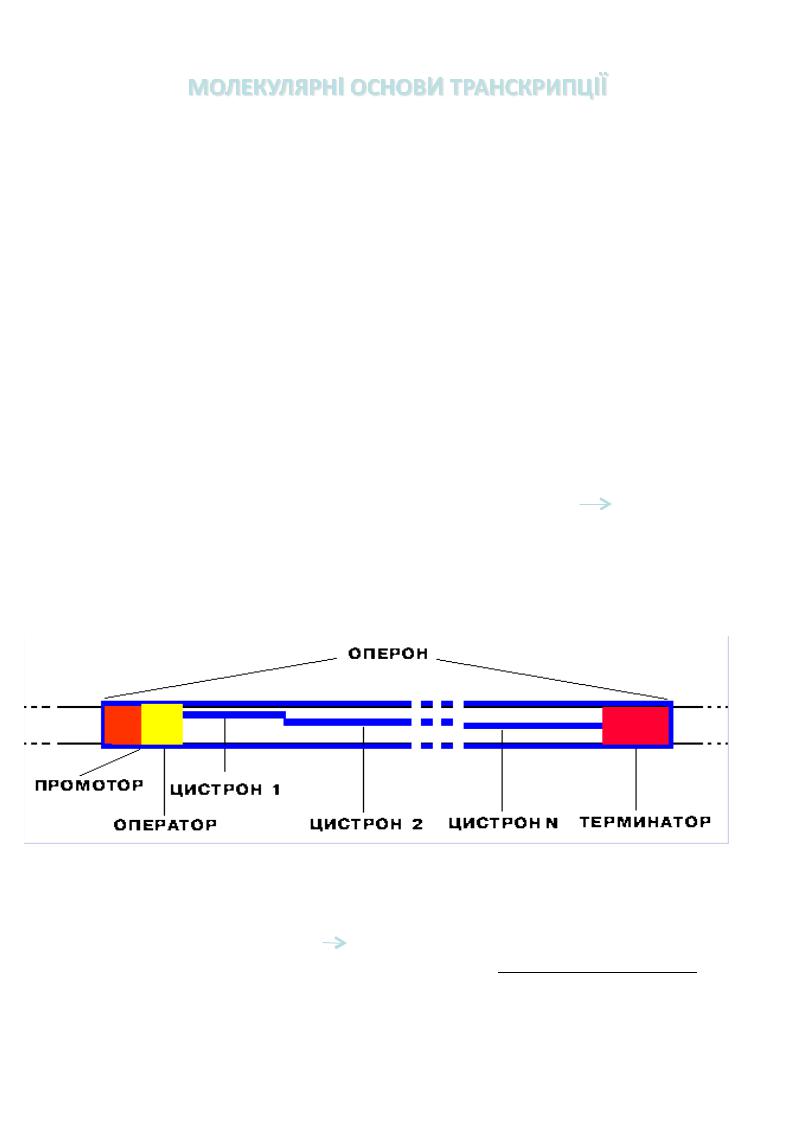
РНК синтезується комплементарно і антипаралельно ланцюгу ДНК, який транскрибується . Подовження ланцюга РНК йде тільки у напрямку5' → 3'. Для початку синтезу РНК фермент не потребує поліабо олігонуклеотидну приманку . Ланцюг ДНК, на якому йде транскрипція , називається кодуючим ,
інший |
некодуючим . Ділянка ДНК, яка підлягає процесу транскрипції , |
|
називається транскриптон (еукаріоти ) або оперон (прокаріоти ) |
(300 – |
|
1011нуклеотидів ) |
|
|
Структура!оперона!
Промотор!:""участок оперона , існує для впізнавання ферментомРНКполімеразою .Послідовність основ по ходу ланцюга ДНК, що знаходиться нижче
за сайтом промотора в напрямку 3’→ 5’, використовується як матриця для синтезу РНК. До промотора може приєднуватиськомплекс цАМФ-БАК- білокактиватор катаболічного гена), що полегшує РНК-полімеразі початок транскрипції
структурних генів.
139
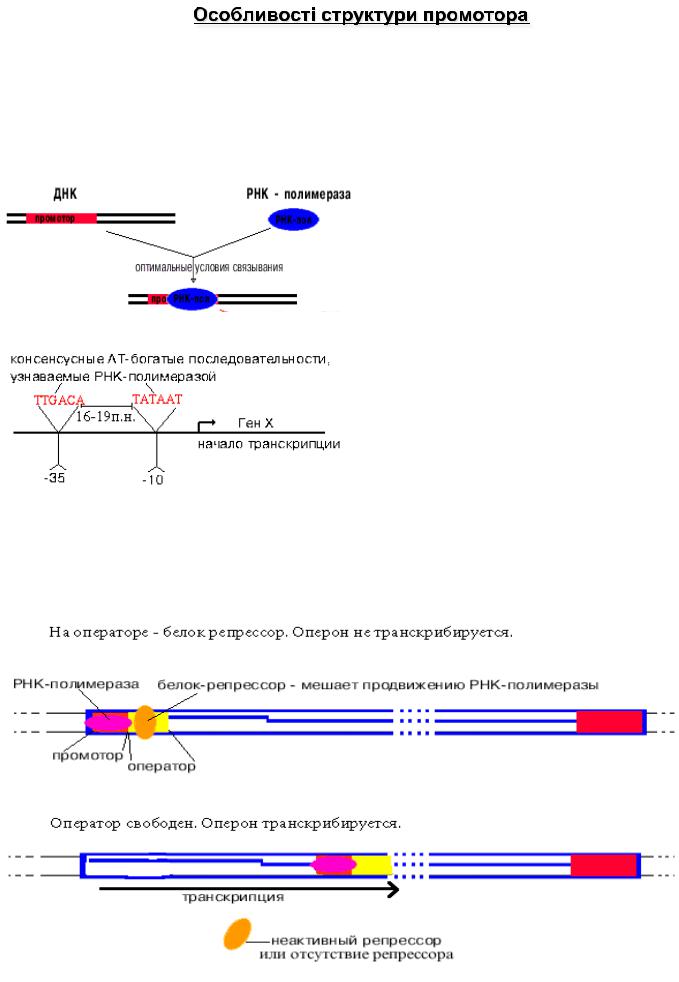
Пізнавання і міцне скріплення відбувається на різних ділянках ДНК. Ці ділянки відрізняються і по первинній , і по вторинній структурі . Шляхом
секвенірування виявили структуру багатьох промоторів . У більшості з них є
|
загальна властивість."" |
|
|
|
РНК-полімераза впізнає промотор , |
||
|
покриваючи 40-60 пар нуклеотидів . В |
||
|
промоторі впізнається взаємне |
||
|
розташування двох |
AT-багатих |
|
|
ділянок. В кожному з них 4-6 пар |
||
|
нуклеотидів . Центри цих ділянок |
||
|
знаходяться в положеннях "-10" і |
||
|
"-35". Принципово важливою є |
||
|
відстань між цими ділянками . Вона |
||
|
коливається від 16 до 19 п.н. Штучне |
||
|
збільшення цієї відстані до 20 п.н. або |
||
|
зменшення його до 15 п.н. призводить |
||
|
до того, що РНК-полімераза не |
||
|
впізнає зіпсований промотор."" |
||
Після промотора в опероні знаходиться |
акцепторна зона (у еукаріот ) або |
||
оператор |
(у прокаріот ), яка служить для скріплення з регуляторами |
||
транскрипції |
(наприклад , підсилювачами |
- енхансерами ). У оператора |
|
диспетчерська функція : він дозволяє або забороняє транскрипцію.
140