

9.1. Содержание экстенсина и оксипролина в клеточных стенках некоторых растительных продуктов (%)
Продукт |
Экстенсии |
Оксипролин |
Картофель |
22,0 |
1,2 |
Морковь |
12,0 |
0,6 |
Свекла |
11,9 |
1,6 |
Петрушка (корень) |
8,8 |
0,4 |
Дыня |
2,8...5,0 |
0,08...0,36 |
ном из кислых полисахаридов (протопектина), играющих роль межклеточного цементирующего вещества, которому иногда сопутствуют протеиновые соединения, а в наиболее старых тканях-- лигнин.
Кроме углеводов в клеточных стенках содержатся азотистые вещества, лигнин, липиды, воска, минеральные вещества.
Из азотистых веществ в клеточных стенках растительной ткани обнаружен структурный белок экстенсии — полимер из группы гликопротеидов, белковая часть которого связана с углеводами — остатками арабинозы и галактозы. Молекулярная масса белковой части таких макромолекул равна 50 000, экстенсии имеет форму жесткого стержня, на 50 % состоит из оксипролина. В клеточной стенке присутствует несколько фракций белка, различающихся содержанием оксипролина.
Экстенсии в некоторых отношениях напоминает белок коллаген, выполняющий аналогичные функции в животных тканях. Содержание экстенсина и оксипролина в клеточных стенках различных овощей и картофеля неодинаково (табл. 9.1). Клеточные стенки картофеля состоят примерно на 1/5 из экстенсина. В клеточных стенках корнеплодов его содержится в 2 раза меньше, чем в клеточных стенках картофеля; в клеточных стенках дыни содержание экстенсина не превышает 5 %.
Содержание оксипролина в клеточных стенках этих растительных продуктов колеблется в зависимости от вида продукта от сотых долей процента до 1,5 % и более.
Содержание в клеточных стенках экстенсина и оксипролина изменяется в процессе хранения овощей. Особенно заметны эти изменения при повреждении ткани овощей. Так, в дынях при повреждении плодов содержание белков в клеточных стенках возрастает в 3...4 раза, а оксипролина — в 5... 10 раз.
Соотношение углеводов и экстенсина в клеточных стенках зависит от вида растительной ткани. Клеточные стенки многих растительных продуктов состоят примерно на 1/3 из целлюлозы, на 1/3 из гемицеллюлоз и на 1/3 из пектиновых веществ и белка. В клеточных стенках томатов между углеводами и белком существует другое соотношение —1:1.
Лигнин — природный полимер сложного строения, формирующий клеточные стенки растений. Играет роль инкрустирующего вещества, скрепляющего волокна целлюлозы и гемицеллюлоз. Ковалентно связан с полисахаридами гемицеллюлоз (ксиланом), с пектиновыми веществами и белком. Содержание лигнина в тканях растений зависит от их вида и степени одревеснения. Значительное количество лигнина содержится в клеточных стенках свеклы, моркови, меньше его накапливается в белокочанной капусте.
В связи с тем что размягчение картофеля, овощей и плодов, происходящее в процессе их тепловой кулинарной обработки, связывают с деструкцией клеточных стенок, представляется целесообразным рассмотреть строение последних.
По современным представлениям, клеточная стенка — это высоко специализированный агрегат, состоящий из различных полимеров (целлюлозы, гемицеллюлоз, пектиновых веществ, белков и др.), структура которых у разных растений закодирована с той же степенью точности, что и структура молекул белков. На рис. 9.4 представлена модель структуры первичной клеточной стенки.
Первичная клеточная стенка состоит из волокон (микрофибрилл) целлюлозы, которые занимают менее 20 % объема гидра-тированной стенки. Располагаясь в клеточных стенках параллельно, целлюлозные волокна с помощью водородных связей образуют мицеллы, которые имеют правильную, почти кристаллическую упаковку. Одна мицелла целлюлозы может отстоять от другой на расстоянии, равном десяти ее диаметрам. Пространство между мицеллами целлюлозы заполнено аморфным основным веществом (матриксом), состоящим из пектиновых веществ, гемицеллюлоз (ксилоглюкан и арбиногалантан) и структурного белка, связанного с тетрасахаридами.
Первичная стенка клетки рассматривается как целая мешкообразная макромолекула, компоненты которой тесно взаимосвязаны. Между мицеллами целлюлозы и ксилоглюканом существуют многочисленные водородные связи. В свою очередь, ксилоглюкан ковалентно связан с боковыми галактановыми цепями
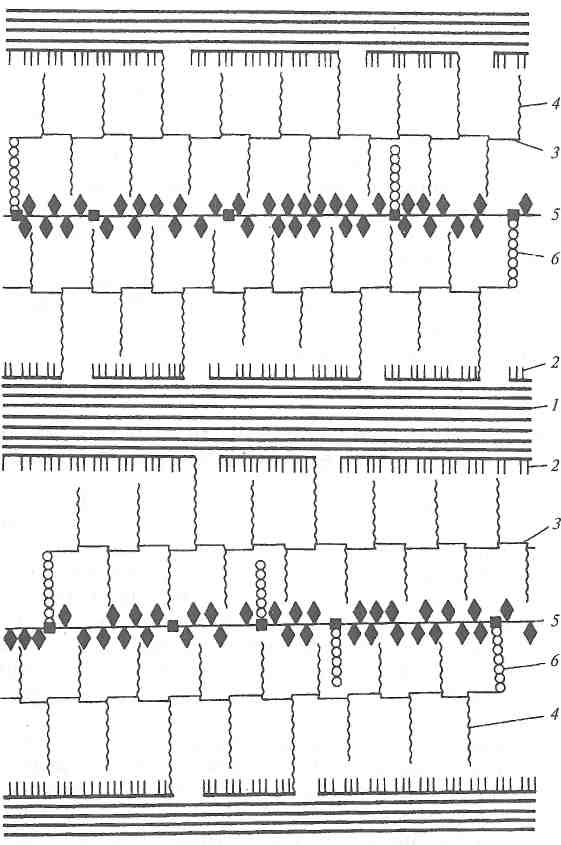
Рис. 9.4. Структура первичной клеточной стенки (по Альберсхейму):
1 — микрофибрилла целлюлозы: 2 — ксилоглюкан; 3 — главные
рамногалактуроновые цепи пектиновых веществ; 4— боковые
галактановые цепи пектиновых веществ; 5 — структурный белок
с арабинозными тетрасахаридами; 6— арабиногалактан
пектиновых веществ, а пектиновые вещества через арабинога-лактан ковалентно связаны со структурным белком.
Учитывая, что клеточные стенки многих овощей и плодов отличаются относительно высоким содержанием двухвалентных катионов, в основном Са и Mg (0,5... 1,0 %), между пектиновыми молекулами, содержащими свободные карбоксильные группы, могут возникать хелатные связи в виде солевых мостиков.
Вероятность образования солевых мостиков и степень этери-фикации полигалактуроновых кислот связаны обратной зависимостью. Солевые мостики способствуют упрочнению клеточных стенок и паренхимной ткани в целом.
Покровные ткани клубней картофеля, корнеплодов и других овощей характеризуются пониженной пищевой ценностью из-за концентрации в них клетчатки и гемицеллюлоз, поэтому при кулинарной обработке картофеля и большинства овощей эти ткани удаляют.
ФИЗИКО-ХИМИЧЕСКИЕ ПРОЦЕССЫ, ПРОИСХОДЯЩИЕ ПРИ КУЛИНАРНОЙ ОБРАБОТКЕ КАРТОФЕЛЯ, ОВОЩЕЙ И ПЛОДОВ
При механической кулинарной обработке картофеля, овощей и плодов (очистка, нарезка, промывание, отжимание сока и др.) частично нарушается целостность их паренхимной ткани, а часть клеток и отдельных клеточных структур разрушается. Это облегчает переход основных пищевых веществ из разрушенных клеток в окружающую среду, а также смешивание содержимого их клеточных органелл. В результате масса продуктов и их пищевая ценность изменяются, возникают ферментативные, окислительные и другие процессы, вызывающие изменение органолептиче-ских показателей (цвета, вкуса, консистенции) продукта.
В начальный период тепловой кулинарной обработки картофеля, овощей и плодов могут активизироваться все содержащиеся в них ферменты, вызывающие те или иные изменения пищевых веществ. На определенном этапе тепловой обработки ферменты инактивируются, цитоплазма и клеточные мембраны вследствие денатурации белков разрушаются, отдельные компоненты клеточного сока и других структурных элементов клетки получают возможность взаимодействовать друг с другом и окружающей средой.
В результате окислительных, гидролитических и других процессов изменяются химический состав продуктов, их структурно-механические характеристики и органолептические показатели.
Представляют интерес такие физико-химические процессы, происходящие в картофеле, овощах и плодах при тепловой кулинарной обработке, которые вызывают изменения механической прочности паренхимной ткани (размягчение), консистенции, массы, содержания основных пищевых веществ, цвета, вкуса и аромата.
Размягчение картофеля, овощей и плодов
Размягчение тканей картофеля, овощей и плодов, как правило, происходит при тепловой кулинарной обработке. Без воздействия теплоты размягчение наблюдается в основном в плодах (яблоки, груши, бананы и др.) и некоторых овощах (томаты) в процессе созревания и хранения технически спелой продукции вследствие процессов, протекающих в них под действием ферментов. Частичное размягчение тканей капусты белокочанной наблюдается при квашении, что связано, по-видимому, как с ферментативными процессами, так и с кислотным гидролизом протопектина, которого в клеточных стенках квашеной капусты содержится в 1,5 раза меньше, чем в свежей.
Подвергнутые тепловой кулинарной обработке картофель, овощи и плоды приобретают более мягкую консистенцию, легче раскусываются, разрезаются и протираются. Степень размягчения картофеля, овощей и плодов в процессе тепловой обработки оценивают по механической прочности1 их тканей. Так, механическая прочность образцов сырого картофеля при испытании их на сжатие составляет около 13 • 105 Па, а вареного — 0,5 ■ 105 Па, образцов сырой свеклы — 29,9 • 105 Па, вареной — 2,9 ■ 105 Па.
Размягчение картофеля, овощей и плодов при тепловой кулинарной обработке связывают с ослаблением связей между клетками, обусловленным частичной деструкцией клеточных стенок.
Деструкция клеточных стенок
Представленные на рис. 9.5 микрофотографии ткани сырой и вареной свеклы свидетельствуют о том, что клеточные стенки последней отличаются более разрыхленной структурой.
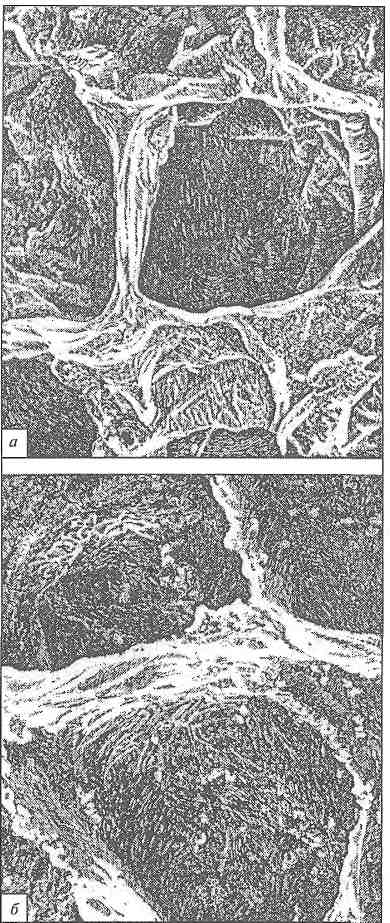
Рис. 9.5. Микрофотографии ткани сырой (а) и вареной (б) свеклы (по Г. М. Харчук)
Однако при доведении овощей и плодов до кулинарной готовности клеточные стенки не разрываются. Более того, клеточные оболочки вареных овощей не разрываются при протирании и раскусывании, так как обладают достаточной прочностью и эластичностью. В этих случаях ткань разрушается по срединным пластинкам, которые подвергаются деструкции в большей степени, чем клеточные оболочки.
Благодаря этому при разжевывании вареного картофеля не ощущается, например, вкус крахмального студня. Клеточные оболочки не разрушаются даже при очень длительной тепловой обработке овощей и плодов, когда может происходить частичная мацерация их тканей (распад на отдельные клетки).
Установлено, что в процессе тепловой кулинарной обработки картофеля, овощей и плодов глубоким изменениям подвергаются нецеллюлозные полисахариды клеточных стенок: пектиновые вещества и гемицеллюлозы, а также структурный белок экстенсии, в результате чего образуются продукты, обладающие различной растворимостью. Именно степень деструкции полисахаридов и растворимость продуктов деструкции обусловливают изменение механической прочности клеточных стенок овощей и плодов при тепловой кулинарной обработке. Изменения целлюлозы в этом случае сводятся главным образом к ее набуханию.
Деструкция протопектина
Известно, что при тепловой кулинарной обработке картофеля, овощей, плодов и других растительных продуктов содержание протопектина в них уменьшается. Так, при доведении овощей до кулинарной готовности содержание протопектина в них может снижаться на 23...60 % (табл. 9.2).