

Тема 2. Структурно-функциональная характеристика нервной ткани. Биоэлектрические явления.
В основе современного представления о структуре и функции ЦНС лежит нейронная теория.
Нервная система построена из двух типов клеток: нервных и глиальных, причем число последних в 8 - 9 раз превышает число нервных. Однако, именно нейроны обеспечивают все многообразие процессов, связанных с передачей и обработкой информации.
Нейрон, нервная клетка, является структурно-функциональной единицей ЦНС. Отдельные нейроны, в отличие от других клеток организма, действующих изолированно, «работают» как единое целое. Их функции состоит в передаче информации (в форме сигналов) от одного участка нервной системы к другому, в обмене информацией между нервной системой и различными участками тела. При этом передающие и принимающие нейроны объединены в нервные сети и цепи.
В нервных клетках происходят сложнейшие процессы обработки информации. С их помощью формируются ответные реакции организма (рефлексы) на внешние и внутренние раздражения.
Нейроны обладают рядом признаков, общих для всех клеток тела. Независимо от своего местонахождения и функций, любой нейрон, как всякая другая клетка, имеет плазматическую мембрану, определяющую границы индивидуальной клетки. Когда нейрон взаимодействует с другими нейронами, или улавливает изменения в локальной среде, он делает это с помощью мембраны и заключенных в ней молекулярных механизмов.
Нейроны, в отличие от других клеток, кроме тела (сомы) снабжены отростками. Многочисленные короткие древовидно разветвленные отростки дендриты служат своеобразными входами нейрона, через которые сигналы поступают в нервную клетку. Они имеют шероховатую поверхность, создаваемую небольшими утолщениями шипиками, словно бусинками, нанизанными на дендрит. Благодаря этому увеличивается поверхность нейрона и максимально повышается сбор информации.
Выходом нейрона является отходящий отросток аксон, который передает нервные импульсы дальше другой нервной клетке или рабочему органу (Рис.1). Аксоны многих нейронов покрыты миелиновой оболочкой. Она образована швановскими клетками, многократно (до 10 и более слоев) «обернутыми» подобно изоляционной ленте вокруг ствола аксона. Однако, муфты швановских клеток, надетые на аксон, не соприкасаются друг с другом. Между ними остаются узкие щели перехваты Ранвье. Только здесь нервное волокно непосредственно соприкасается с внеклеточной жидкостью. Поэтому, в нервной системе волна распространяющегося нервного импульса бежит не плавно, а движется скачками от одного перехвата к другому, что весьма ускоряет процесс распространения импульса.
Что же касается начальной части аксона в месте выхода его из тела клетки (область «аксонного холмика»), то она лишена миелиновой оболочки. Мембране этой немиелиновой части нейрона так называемого начального сегмента обладает высокой возбудимостью. Поэтому ее называют пусковой зоной, так как именно отсюда начинается возбуждение нейрона.
Нет необходимости говорить о том, что даже для внутримозговых связей нужны очень длинные отростки, не говоря уже об аксонах, выходящих за пределы ЦНС к мышцам, железам, внутренним органам. Собранные в пучки, они образуют нервы.
Если нейрон образует выходные связи с большим членом других клеток, то его аксон может многократно ветвиться, чтобы сигналы могли дойти до каждой из них, количество таких разветвлений колеблется от 1000 до 10000 и более. Кроме того, аксон способен отдавать дополнительные ветви - коллатерали, по которым возбуждение уходит далеко в сторону от магистрального пути.
Форма нервной клетки, ее размеры и расположение отростков разнообразны и зависят от функционального назначения нейрона.
Каждый отдельный нейрон уникален и неравноценен себе подобным, в отличие от других клеток организма. Величина нейронов весьма вариабельна: самые крупные в десятки и сотни раз больше самых мелких.
На вопрос, сколько же нейронов содержит мозг человека и высокоорганизованных животных никто точно ответить не может, но считается, что их количество измеряется приблизительно десятками миллиардов.
Классификация нейронов
Современная нейробиология предлагает два принципа классификации нейронов - по форме, строению и выполняемым ими функциям:
I. Классификация нейронов по форме: звездчатые, овальные, округлые, пирамидальные, клетки Беца (мозг).
II. Классификация нейронов по строению
-
Большинство нейронов состоят из тела, нескольких отходящих от него дендритов и одного аксона - мультиполярные нейроны:
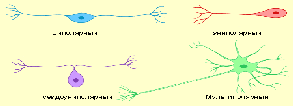
Рис. Виды нервных клеток.
2. Нейроны, состоящие из тела, аксона и одного дендрита, называются биполярными.
3. Униполярными называются нейроны, воспринимающие возбуждение за счёт синапсов, расположенных на теле клетки, и передающие его по единственному отростку - аксону. У человека такие нейроны обнаружены только в чувствительном ядре тройничного нерва на уровне среднего мозга. Существуют нейроны, которые по своей структуре являются униполярными, но функционально они относятся к биполярным клеткам. От тела этих клеток отходит один отросток (аксон), но его проксимальная часть Т-образно разветвляется на два волокна: афферентное и эфферентное. Такие нейроны называются псевдоуниполярными; они расположены в спинномозговых ганглиях (ганглиях задних корешков) и в чувствительных ганглиях черепно-мозговых нервов. Уникальность этих клеток заключается в том, что по миелинизированным афферентным отросткам импульсы проходят намного быстрее, чем по обычным дендритам, не покрытым миелиновой оболочкой.
4
III. Нейроны делятся на возбуждающие и тормозные. Отсюда следует, что нейроны «специализирующиеся» на процессах возбуждения, называются возбуждающими, а на процессах торможения - тормозными.
IV. В зависимости от выполняемых функций обычно выделяют нейроны (рис ):
Афферентные (чувствительные, центростремительные, сенсорные) - передают импульсы (информацию) от рецепторов в ЦНС. Тела этих нейронов расположены вне ЦНС - в спинномозговых или черепно-мозговых ганглиях (рядом с головным и спинным мозгом). Афферентный нейрон имеет псевдоуниполярную форму, т.е. оба его отростка выходят из одного полюса клетки. Один из его отростков направляется на периферию, где заканчивается рецептором, а другой - в ЦНС. К афферентным нейронам также относятся нервные клетки, аксоны которых составляют восходящие пути головного и спинного мозга.
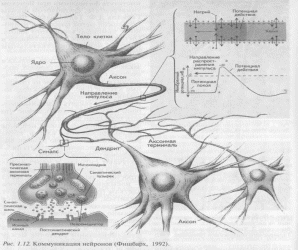
Рис Коммуникация нейрона
2. Эфферентные (эффекторные, двигательные и вегетативные) нейроны работают в центробежном режиме, т.е. они связаны с передачей нисходящих импульсов от вышерасположенных этажей нервной системы к нижерасположенным. Например, от коры к спинному мозгу, или от спинного мозга к рабочим органам. Для эфферентных нейронов характерна разветвленная сеть дендритов и один длинный аксон. Необходимо отметить, что количество эфферентных нейронов в 4-5 раз меньше афферентных.
3. Вставочные (промежуточные, интернейроны, сочетательные, ассоциативные) как правило, более мелкие клетки, осуществляющие связь между различными нейронами (в частности, афферентными и эфферентными). Они передают нервные импульсы в различных направлениях (горизонтальном, вертикальном) по ЦНС. Благодаря многочисленным разветвлениям аксона промежуточные нейроны могут одновременно возбуждать большое число других нейронов. В ЦНС преобладают промежуточные нейроны. Особое место занимают модуляторные нейроны, которые самостоятельно не запускают каких-либо реакций, но могут изменять уровень активности нервных центров, модулируя, таким образом, их реактивность.
4. Секреторные нейроны вырабатывают различные гормоны, выделяющиеся в кровь и осуществляющие гуморальную регуляцию работы различных органов и систем (нейроны гипоталамуса и гипофиза).
Из цепи функционально специализированных нейронов строятся рефлекторные дуги: простые (двухнейронные, моносинаптические) и очень сложные (полисинаптические). Связь между нейронами осуществляется посредством синапсов, чаще всего - химических. Нейрон, передающий информацию через синапс, называется пресинаптическим; получающий информацию нейрон называется постсинаптическим.
Глиальные клетки: их разновидности и функции. Нейроны в нервной системе окружены опорными и вспомогательными клетками, которые называются глиальными. Количество глиальных клеток в ЦНС в 5-10 раз превышает количество нейронов.
Клетки нейроглии плотно окружают всю сосудистую капиллярную сеть в мозговой ткани. Свободной остается лишь незначительная часть поверхности сосудов (около 15%). Выросты глиальных клеток могут располагаться с одной стороны на нейроне, с другой - на кровеносных сосудах. Это указывает на их важное значение в передаче питательных веществ и кислорода из крови в нервную клетку. Доказано, что нейроглия активно участвует в функционировании нейрона. В случае необходимости, компенсируется доставка питательных веществ и кислорода к активно «работающим» нейронам.
Клеткам нейроглии иногда приписывают также функцию своеобразной гидродинамической подушки, предохраняющей чувствительные и нежные образования нейронов от различных физических воздействий.
В последнее время появились сообщения об участии этих клеток в условно рефлекторной деятельности мозга и механизмах памяти.
Таким образом, система «нейрон – нейроглия» постоянно находится в состоянии гибкого ритмически колеблющегося равновесия. Но в этих процессах, происходящих в мозге, безусловно господствуют нейроны, и, пользуясь своим положением, тянут из нейроглии все, что ей нужно.
Глиальные клетки бывают нескольких типов. Три типа клеток - олигодендроциты, астроциты и эпендимные клетки - относятся к нейроглиальным клеткам, то есть имеют общее происхождение с нейронами, но, в отличие от них, способны к регенерации. Клетки микроглии - являются макрофагами, мигрировавшими из кровотока в ткани мозга.
4
1
2
1. Олигодендроциты обнаружены в белом веществе головного и спинного мозга, в периферических нервах.
5
3
Аналогичные клетки в периферической нервной системе называются шванновскими клетками (клетками-спутниками, клетками-сателлитами).
Олигодендроциты образуют отростки, которые покрывают и изолируют нервные клетки и волокна. Олигодендроциты заключают их в складки своей наружной мембраны. При этом мембрана отростков олигодендроцитов, сопровождающих нервные волокна А и В типа, как бы накручивается вокруг соответствующего фрагмента каждого аксона. В результате эти клетки покрывают своей цитоплазматической мембраной ствол аксона в несколько слоёв с небольшими межклеточными промежутками между ними, называемыми перехватами Ранвье. После накручивания своих отростков вокруг аксона олигодендроцит начинает формировать специализированную клеточную оболочку, включая в её состав в качестве основного структурного белка миелин. Образовавшийся многослойный мембранный комплекс называется миелиновой оболочкой. Миелин, таким образом, образован мембранными белками и липидами, которые обусловливают белый цвет участков нервной ткани, состоящих преимущественно из миелинизированных волокон (белое вещество головного и спинного мозга).
В периферической нервной системе миелинизацию осуществляют шванновские глиальные клетки. Шванновские клетки, в отличие от олигодендроцитов ЦНС, отростков не образуют; каждая из них как бы обвёртывает собой участок аксона, образуя вместе с другими шванновскими клетками его миелиновую оболочку. Между соседними шванновскими клетками остаются небольшие свободные участки аксонов (перехваты Ранвье), по которым распространяются нервные импульсы.
2
Олигодендроциты и шванновские клетки, заключающие в своих складках тела нейронов и немиелинизированные нервные волокна (С тип), могут одновременно образовывать изолирующие «футляры» для нескольких нейронов и их отростков.

Рис. Формы глиальных клеток
2. Астроциты имеют звёздчатую форму (рис.3). Некоторые из них снабжены тонкими цитоплазматическими отростками (фибриллярные астроциты), а часть - плотными отростками (протоплазматические астроциты).
Находятся астроциты преимущественно в сером и белом веществе головного и спинного мозга. Отростки заполняют пространства между сосудистыми стенками и нейронами. Это поддерживающие и изолирующие клетки, обеспечивающие транспорт веществ из кровеносных капилляров к нервным клеткам.
Астроциты обеспечивают нейроны питательными веществами, поступающими по сосудам (трофическая функция) и одновременно участвуют в формировании гематоэнцефалического барьера (ГЭБ), препятствующего поступлению из крови вредных веществ (защитная функция).
3. Эпендимные клетки образуют непрерывную выстилку стенок желудочков мозга и центрального канала спинного мозга. Эти клетки глубоко проникают в подлежащую нервную ткань. На апикальной поверхности эпендимных клеток, которая обращена в просвет желудочков или спинномозгового канала, расположено большое количество микропиноцитозных пузырьков и микроворсинок. Эпендимные клетки выполняют транспортную и секреторную функцию, принимая участие в образовании спинномозговой жидкости.
4. Микроглия представлена мелкими клетками с множеством отростков. Клетки микроглии выполняют в ЦНС фагоцитарную функцию, удаляя погибшие нервные и глиальные клетки, вирусы и бактерии. Выполняет роль барьера между веществом мозга и омывающей его спинномозговой жидкостью; регулирует секрецию и состав спинномозговой жидкости.
развиваются в процессе эмбриогенеза из нервной трубки. Сложное взаимное расположение нейронов и клеток нейроглии обусловливает вид нервной ткани на разрезе, когда участки серого вещества чередуются с участками белого вещества. В спинном мозгу серое вещество занимает центральную позицию и имеет вид бабочки. В мозжечке и в полушариях большого мозга серое вещество образует ядра (участки скоплений нейронов при относительно небольшом количестве глиальных клеток, отделённые друг от друга белым веществом) и кору мозжечка и больших полушарий. Серое вещество коры большого мозга состоит из 6 слоёв нервных клеток, а в коре мозжечка таких слоёв три.
Биоэлектрические потенциалы в возбудимых тканях. К концу ХIХ и в начале ХХ стало известно, что электрические потенциалы образуются во всех живых тканях, а возникновение и распространение возбуждения связано с изменением электрического заряда на поверхности клеточной мембраны и внутри клетки.
В 1902 году Ю.Бернштейном была предложена мембранная теория возникновения электрических потенциалов живых тканей. Суть ее – потенциалы возникают за счет разности зарядов внутри и снаружи клетки.
теории натрий-калиевого насоса.
Было установлено, что величина внутриклеточного электрического потенциала у нервных клеток и волокон в покое составляет 70 милливольт (мВ). Эту величину называют мембранным потенциалом покоя (МПП).
Природа поляризации клеточной мембраны сегодня в основном определена. Мембрана аксона как и других клеток, тонкая, но достаточно прочная. Состоит мембрана из липидов, белков, мукополисахаридов. Мембрана легко проницаема для жирорастворимых веществ, молекулы которых легко проникают через липидный матрикс.
Крупные водорастворимые молекулы, в том числе анионы органических кислот практически не проникают мембрану и покидают клетку лишь путем экзоцитоза.
В то же время в мембране нервного волокна существуют каналы, проницаемые для воды, для малых молекул водорастворимых веществ и для малых ионов. Особое значение имеют каналы, проницаемые для ионов Nа +, К+, Cl‾, С²+. В нервной мембране обнаружены специфические натриевые, калиевые, хлорные и кальциевые каналы, то есть каналы избирательно пропускающие названные ионы. Эти каналы могут быть открытыми или закрытыми, то есть обладают воротными механизмами. В покое практически все натриевые каналы мембраны клетки закрыты, а большинство калиевых каналов открыты.
Для возникновения МПП, т.е. мембранного потенциала покоя, важно отличие ионного состава плазмы клетки от ионного состава внешней среды. В цитоплазме меньше ионов Nа+ и больше К+ и несравненно больше органических анионов. Катионные различия объясняются результатами работы так называемого калиево-натриевого насоса мембраны, непрерывно откачивающего Nа+ из клетки в обмен на К+. Движение ионов через мембрану называется активным ионным транспортом (рис..)
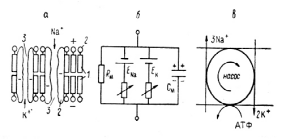
Рис.3. 1. Упрощенные схемы строения плазматической мембраны и ее механизмов, ответственных за формирование потенциала покоя и потенциала действия.
а - схема строения и б - эквивалентная электрическая схема мембран; в - схема работы ионного насоса. 1 - билипидный матрикс, 2- белковые молекулы; 3 - открытые катионные каналы.
Движение ионов, по сути, главный механизм формирования МПП. Но в образовании МПП участвует добавочный механизм – прямой электрогенный эффект натрий-калиевого насоса. Этот эффект зависит от поляризации мембраны, возникающей за счет неравенства числа ионов Nа+ и К+
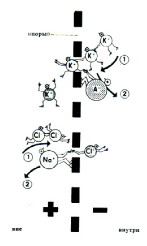
Рис.3.2. Возникновение электрохимического равновесия на мембране.
Диффузионное давление (1) в равновесии с противоположной электростатической силой (2), стремящейся удержать вместе ионы с противоположными зарядами.
Разность зарядов между наружной и внутренней поверхностью клеточной мембраны в состоянии физиологического покоя клетки называется потенциалом покоя. В самой мембране МПП проявляется как электрическое поле значительной напряженности, которое обеспечивает закрытое и открытое состояние ворот натриевые каналов, обеспечивающих состояние покоя и готовности к возбуждению.
Потенциал действия или потенциал возбуждения нервных клеток возникает в ответ на достаточное по силе раздражение. Это очень быстрый, кратковременный электрический процесс, который проявляется, в первую очередь, изменением электрического состояния клеточной мембраны. Раздражение способствует перезарядке, реверсии, т.е. заряд мембраны уменьшается от нуля и приобретает отрицательное значение: внутренняя сторона мембраны заряжается положительно, наружная – отрицательно (рис.). Затем вновь восстанавливается первоначальный заряд. Колебания потенциала, возникающее в результате кратковременной перезарядке мембраны и последующего восстановления ее исходного заряда, называется потенциалом действия. В потенциале действия различают пик (длительность 0,5 – 0,8 мс) и следовые потенциалы (рис. )
Потенциал действия имеет несколько фаз:
1Фаза нарастания - потенциал действия начинается быстрым сдвигом в положительном направлении. В это время клеточная мембрана поляризована.
2. Овершут - деполяризация переходит за нулевую линию, и мембранный потенциал становится положительным.
3. Реполяризация - фаза, в течение которой восстанавливается исходный потенциал мембраны в покое. Последний участок фазы реполяризации для некоторых видов потенциала действия бывает замедлен.
4. Гиперполяризационный потенциал - кривая деполяризации быстро пересекает уровень потенциала покоя, так что на некоторое время потенциал становится более отрицательным, чем потенциал покоя.
5. Деполяризационным следовым потенциалом - примерно через 1 мс после начала потенциала действия наблюдается отчетливый перегиб кривой реполяризации.
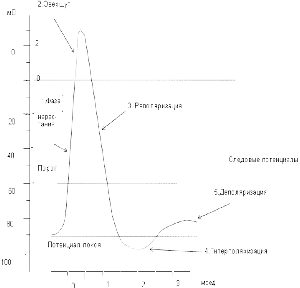 Рис.
Фазы потенциала действия.
Рис.
Фазы потенциала действия.
Современная мембранная теория объясняет механизм возникновения потенциала действия прохождением потоков ионов калия и натрия через каналы. Когда деполяризация достигает критической величины, за доли секунды, одновременно открываются максимальное количество натриевых каналов, проницаемость мембраны для ионов натрия увеличивается по сравнению с покоем в 500 раз. Ионы натрия устремляются внутрь клетки, унося с наружной поверхности мембраны положительный заряд, благодаря чему положительно заряженной становится внутренняя поверхность мембраны. Наружная же поверхность мембраны становится отрицательно заряженной.
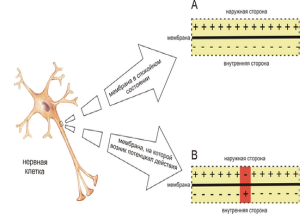
Рис. Ионы натрия устремляются внутрь клетки, положительно заряжают внутреннюю поверхность мембраны. Наружная поверхность мембраны становится отрицательно заряженной.
Но такое положение длится всего 0,5 – 1,5 мс. Количество открытых натриевых каналов уменьшается, проницаемость для ионов натрия уменьшается до исходного уровня, этот процесс называется инактивацией механизма натриевой проницаемости. Но во время инактивации механизма натриевой проницаемости увеличивается проницаемость мембраны для ионов калия. Выходя из клетки, ионы калия выносят положительный заряд, чем восстанавливают исходную разность потенциалов и мембрана опять приобретает снаружи положительный заряд, изнутри – отрицательный.
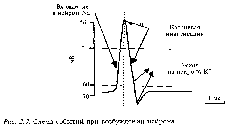
Рис. Схема событий при возбуждении нейрона.
Потенциал действия, возникнув в одном участке нервной клетки, нервного или мышечного волокна, быстро распространяется по всей ее поверхности. Потенциал действия по силе превышает в 5-10 раз силу раздражения, способную вызвать возбуждение, это необходимо для возбуждения участка клетки, находящегося в покое. Между возбужденным участком и находящимися в состоянии покоя, возникает разность потенциалов, приводящая к возникновению круговых токов.
Таким образом, потенциал покоя определяется ионами калия, а потенциал действия зависит от ионов натрия.
Скорость проведения возбуждения в волокнах скелетных мышц достигает 12-15 м/с, гладких мышц – 2-15, в безмякотных нервных волокнах – 0,5 – 3, в мякотных – 70-120 м/с.
Основные характеристики нервных тканей. Для любой живой клетки характерны следующие свойства: раздражимость, возбудимость, функциональная подвижность (лабильность).
Ι. Раздражимость – общее свойство любой живой ткани, клетки реагировать на раздражение изменением обмена веществ и энергии. Следует учитывать, что изменения носят местный характер, т.е. реагирует только участок ткани, который подвергается раздражению.
Раздражители можно условно разделить на три группы:
-
Физические механические, температурные, электрические, световые, невидимые глазом инфракрасное и ультрафиолетовое излучения, радиоактивное излучение.
-
Химические – кислоты, щелочи, яды.
-
Биологические – животные, растения, микробы и вирусы, паразиты высших млекопитающих
Кроме того, раздражители можно разделить на внешние (различные формы раздражения окружающей организм материи) и внутренние (изменение химического состава крови, лимфы, тканевой, спинномозговой жидкости).
Раздражители отличаются также и по силе воздействия.
Естественные раздражители, действующие на данную ткань в природных условия, называются адекватными. Изменения внешней или внутренней среды, являющиеся раздражителями, к восприятию которых не готовы все рецепторы или какой-либо, называются неадекватными. К последней группе относятся физические, механические, электрические раздражители.
ΙΙ. Возбудимость – это свойство нервной клетки отвечать на действие раздражителей изменениями ионной проницаемости мембраны и генерировать потенциал действия.
Минимальная сила (энергия) раздражителя, необходимая для раздражения клетки называется пороговой (порогом). Раздражители, сила которых меньше пороговой, называются подпороговыми. Раздражители, более сильные, чем пороговые – сверхпороговые. Возникнув в точке действия раздражителя, возбуждение способно охватывать всю клетку. Это свойство важно для нервной системы, особенно для ее проводящих путей. При отсутствии достаточных раздражителей в клетке наблюдается состояние физиологического покоя при условии текущего метаболизма. Реакции клетки под воздействием внешних факторов или других клеток организма, выражающиеся в снижении метаболизма, роста, возбудимости, называются торможением.
Для возникновения состояния возбуждения необходимы определенные условия:
а) сила действующего раздражителя должна быть не ниже пороговой;
б) достаточное время действия раздражителя (полезное время). Чем сильнее раздражитель, тем меньше времени потребуется для возбуждения ткани.
в) действие раздражителя с большой силой нарастания.
Изменение возбудимости нервной ткани при возбуждении. Для возбудимости тканей характерны фазовые изменения в зависимости от возникновения и развития импульсов. Во время деполяризации мембраны, при действии возбудителя, и появлении местного возбуждения происходит кратковременное повышение возбудимости. Когда местное возбуждение переходит в распространяющееся, возникает потенциал действия, а возбудимость ткани резко снижается. Возникает состояние, когда ткань после раздражения временно не реагирует на повторное раздражение любой силы – абсолютная рефрактерность (совпадает с пиком потенциала действия) (рис ).

Рис. Распространение возбуждения по немиелинизированному нервному волокну: после прохождения потенциала действия в нервном волокне возникает зона невозбудимости, или рефрактерности.
Период восстановления ткани после абсолютной рефрактерности, когда возбудимость еще понижена называется относительной рефрактерности (совпадает с фазой реполяризации и переходом его в следовую поляризацию).
Затем наступает следующий период – фаза экзальтации – период повышенной возбудимости, совпадающий с периодом следовой поляризации. Со следовой гиперполяризацией совпадает фаза субнормальности, во время которой возбудимость ткани несколько снижена по сравнению с величиной возбудимости в состоянии физиологического покоя.
Очевидно, что фазы изменения возбудимости определены развитием и изменением потенциала действия в живой ткани. Изменение возбудимости зависит от действия натрий-калиевого насоса.
III. Важнейшим физиологическим свойством нервной ткани является функциональная подвижность или лабильность - скорость, с которой в ткани возникает и успевает закончиться полный период отдельного импульса возбуждения. Чем выше уровень процессов в тканях, тем быстрее процессы, составляющие отдельный импульс возбуждения и, следовательно, тем больше отдельных импульсов может образоваться в ткани за определенный отрезок времени. Максимальное число импульсов возбуждения, возникающее за 1 сек. в ответ на такое же число раздражений, называется мерой лабильности. Высокая лабильность характерна для мякотных соматических нервов, меньшая – для безмякотных нервов.
Самый частый ритм раздражений, на которые возбудимая ткань отвечает таким же частым ритмом возбуждения, отражает функциональное состояние ткани или лабильность. Лабильность может изменяться под действием разных условий, в частности зависеть от обмена веществ в ткани. Понижение и повышение физиологической лабильности в связи с деятельностью, проявляющееся более высоким или низким ритмом возбуждения названо усвоением ритма.
Наиболее выгодные в рабочем отношении частоты ритмов раздражения называют оптимум. При этом ритме раздражений новый импульс возникает в момент фазы экзальтации предшествующего импульса, в результате мышца сокращается максимально.
Пессимум ритма раздражения возникает, когда частота раздражений настолько высока, что превышает меру лабильности. Это вызвано тем, что раздражение действует в фазу абсолютной рефрактерности предыдущего импульса. Фаза рефрактерности может углубиться, лабильность будет продолжать снижаться и наступит торможение. Перечисленные явления характерны не только для двигательных реакций, но и для условно-рефлекторных, для секреторных реакций, деятельности сердечно-сосудистой системы.
Парабиоз, его стадии. Каким образом возбуждение переходит в торможение Н.Е.Введенский изложил в классическом труде «Возбуждение, торможение и наркоз».
При действии различных веществ, отличающихся по силе и частоте раздражения, мышца начинает отвечать одинаковыми сокращениями. Эту стадию Введенский назвал уравнительной или трансформирующей.
Следующая стадия сопровождалась сильными сокращениями мышцы при слабых или редких раздражениях, а при действии сильных раздражителей, наоборот, не сокращалась или почти не реагировала. Особенности стадии послужили тому, что ее назвали парадоксальной.
Третья стадия функциональных сдвигов сопровождалась состоянием, когда при воздействии на нерв раздражителя любой силы и частоты – мышца не сокращалась, развилось торможение. Стадия торможения заканчивается состоянием, при котором возбудимость и проводимость, как проявления жизни, отсутствуют, была названа Введенским парабиозом, а последовательные функциональные изменения - стадиями парабиотического процесса. При удалении раздражающих веществ с нерва парабиоз прекращается, функциональные свойства участка альтернирующей ткани восстанавливаются, проходя те же стадии в обратном порядке. Если воздействие раздражителей длительное, нерв отмирает.
Возбуждение и торможение в центральной нервной системе
Сенсорные рецепторы. Импульсы и ЦНС поступают от рецепторов организма.
Различают эффекторные и сенсорные рецепторы.
Эффекторные рецепторы представляют собой белковые структуры клеточных мембран, а также цитоплазмы и ядра, активируются химическими соединениями (медиаторами, гормонами), что запускает ответные реакции клетки.
Эффекторные рецепторы воспринимают раздражители внутренней и внешней среды организма с помощью трансформации энергии раздражения в нервный импульс, раздражителями являются изменение температуры, прикосновении, давление, и т.д.
Основное физиологическое значение сенсорных рецепторов состоит в обеспечении поступления и ЦНС информации о состоянии внешней и внутренней среды, что обеспечивает регуляцию функций внутренних органов и организацию взаимодействия организма и окружающей среды.
Классификация сенсорных рецепторов.
1. По структурно-функциональной организации различают первичные и вторичные рецепторы.
Первичные рецепторы представляют собой чувствительные окончания дендрита афферентного нейрона, тело которого локализуется в спинномозговых ганглиях вегетативных и черепных нервов. К первичным рецепторам относятся тактильные, болевые, температурные, проприорецепторы, обонятельные рецепторы, все рецепторы внутренних органов.
Вторичные рецепторы имеют специальную клетку, синаптически связанную с окончанием дендрита афферентного нейрона. К вторичным рецепторам относятся вестибулярные, слуховые, фоторецепторы и вкусовые рецепторы. Во вторичных рецепторах рецепторный потенциал формируется в рецепторной клетке, а потенциал действия – в окончании дендрита сенсорного нейрона.
-
С психофизиологической точки зрения рецепторы подразделяются в соответствии с органами чувств и формируемыми ощущениями на зрительные, слуховые, вкусовые, обонятельные и тактильные.
-
В зависимости от вида воспринимаемого раздражителя выделяют пять типов рецепторов:
а) механорецепторы расположены в коже, внутренних органах, сосудах, слуховой и вестибулярной системах, опорно-двигательном аппарате;
б) хеморецепторы - локализуются в слизистой оболочке носа, языка, каротидном и аортальном тельцах, продолговатом мозге и гипоталамусе;
в) терморецепторы (тепловые и холодовые) находятся в коже, сосудах, внутренних органах, гипоталамусе, продолговатом, спинном и среднем мозге;
г) фоторецепторы сетчатки глаза;
д) болевые рецепторы (ноцицепторы), раздражителями которых являются механические, термические и химические (гистамин, брадикинин, К+ , Na+ и др.) факторы, локализуются в коже, мышцах, внутренних органах, сосудах, дентине.
-
По расположению в организме различают экстеро - и интероцепторы. К экстерорецепторам относятся рецепторы кожи, видимых слизистых оболочек и органов чувств: зрительные, слуховые, вкусовые, обонятельные, тактильные, кожные болевые и температурные.
-
К интерорецепторам относят рецепторы внутренних органов, рецепторы опорно-двигательного аппарата (проприорецепторы) и вестибулорецепторы.
-
Выделяют также мономодальные рецепторы, для которых имеется только один адекватный раздражитель (зрительные, слуховые, вкусовые, обонятельные, хеморецепторы каротидного синуса) и полимодальные рецепторы, воспринимающие несколько адекватных раздражителей. Например, ирритантные рецепторы легких, воспринимающие механические (частицы пыли) и химические раздражители во вдыхаемом воздухе (пахучие вещества).
-
Согласно скорости адаптации рецепторы делят на три группы: быстро адаптирующиеся, или фазные (рецепторы вибрации - тельца Пачини, прикосновения - тельца Мейснера); медленно адаптирующиеся, или тонические (вестибулорецепторы, проприорецепторы, рецепторы растяжения легких, часть болевых рецепторов); смешанные, или фазнотонические, адаптирующиеся со средней скоростью (терморецепторы кожи, фоторецепторы сетчатки).
Механизм возбуждения рецепторов. При действии на рецептор адекватного раздражителя (к которому он эволюционно приспособлен), который может вызывать конфирмационные изменения воспринимающих структур (активацию рецепторного белка), формируется рецепторный потенциал (РП).
В рецепторах (кроме фоторецепторов) энергия раздражителя после ее преобразования и усиления приводит к открыванию ионных каналов и перемещению ионов, среди которых основную роль играет движение Na+ в клетку. Это приводит к деполяризации мембраны рецептора. Полагают, что в механорецепторах растяжение мембраны ведет к расширению каналов. Рецепторный потенциал является локальным, он может распространяться только электротонически на небольшие расстояния - до 3 мм.
Возникновение ПД в первичных и вторичных рецепторах происходит по-разному.
В первичном рецепторе рецепторная зона является частью афферентного нейрона - окончанием его дендрита. Она прилежит к рецептору. Возникший РП, распространяясь электротонически, вызывает деполяризацию нервного окончания и возникновение ПД. В миелиновых волокнах ПД возникает в ближайших перехватах Ранвье, т.е. в участках, имеющих достаточную концентрацию потенциала зависимых натриевых и калиевых каналов, при коротких дендритах, например в обонятельных клетках - в аксонном холмике. При достижении деполяризации мембраны критического уровня происходит генерация ПД.
Во вторичных рецепторах РП возникает в рецепторной клетке, синаптически связанной с окончанием дендрита афферентного нейрона. Рецепторный потенциал обеспечивает выделение рецепторной клеткой медиатора в синаптическую щель. Под влиянием медиатора на постсинаптической мембране возникает генераторный потенциал, обеспечивающий возникновение ПД в нервном окончании вблизи постсинаптической мембраны. Генераторный потенциал, как и рецепторный, является локальным потенциалом.
Свойства рецепторов:
Высокая возбудимость - для возбуждения фоторецептора сетчатки достаточно одного кванта света, для обонятельного рецептора - одной молекулы пахучего вещества.
Адаптация - уменьшение возбудимости рецепторов при длительном действии раздражителя (только темновая адаптация фоторецепторов приводит к повышению их возбудимости). Адаптация рецепторов выражается в снижении амплитуды РП и, как следствие, в уменьшении частоты импульсации в афферентном волокне.
Спонтанная активность - способность возбуждаться без действия раздражителя, присуща проприорецепторам, фоно-, фото-, вестибуло-, термо-, хеморецепторам.
Кодирование силы раздражителя в рецепторе и афферентном нейроне. Кодирование - преобразование информации в условную форму (код), удобную для передачи по каналу связи. Сила раздражителя в рецепторе кодируется величиной амплитуды РП, который относится к градуальным потенциалам, развивающимся по закону силовых отношений: с увеличением силы стимула возрастает РП, с уменьшением силы стимула - снижается. Следовательно, амплитуда РП зависит в определенных пределах от силы действующего раздражителя.
Во многих рецепторах имеется зависимость между амплитудой РП и силой раздражителя, которая основана на том, что мембранный потенциал изменяется пропорционально ионной проницаемости мембраны.
В некоторых рецепторах между силой раздражителя и амплитудой РП имеется линейная зависимость (механорецепторы внутренних органов) и S-образная зависимость (тельца Пачини).
Амплитуда РП может изменятся плавно, непрерывно, и если деполяризуются до критического уровня участки мембраны афферентного нервного волокна, в котором возможна генерация потенциала действия, то РП перекодирует в импульсную активность нейрона. Частота импульсации афферентного волокна приблизительно пропорциональна амплитуде РП и, следовательно, силе раздражителя. Однако линейность, сохраняемая в пределах лабильности нервного волокна нарушается, если фаза абсолютной рефрактерности ограничмвает частоту импульсации в нем. Возникшее возбуждение под влиянием рецепторного (в первичных рецепторах) или генераторного (во вторичных рецепторах) потенциала движется по афферентным нервным волокнам в ЦНС. В самой ЦНС возбуждение по нервным волокнам (аксонам нейронов) распространяется между нейронами.
Физиология нервного волокна. Классификация нервных волокон. В зависимости от морфо-функциональных особенностей нервные волокна подразделяются на два типа: миелиновые и безмиелиновые. Оболочку безмиелиновых волокон образуют шванновские клетки. Оболочку миелиновых волокон в периферической нервной системе формируют шванновские клетки, а в ЦНС – олигодендроциты. Через равные промежутки, прерываясь, миелиновая оболочка образует перехваты Ранвье (рис. ).

Рис. Образование миелинового чехла на аксоне.
1 – наматывание слоев миелина; 2 – аксон; 3 - олигодендроцит; 4 - перехват Ранвье; 5 – слой миелиновой оболочки
Согласно структурно-функциональным свойствам нервных волокон, выделено три группы нервных волокон: А, В и С, которые разделили по степени выраженности миелиновой оболочки и степени распространения возбуждения.
-
Волокна типа А имеют хорошо выраженную миелиновую оболочку, диаметром 20 мкм, скорость проведения нервного импульса 25-100 м/сек. К ним относятся: моторные волокна скелетной мускулатуры, афферентные нервные волокна, отходящие от рецепторов, воспринимающих давление.
-
Волокна типа В – миелиновая оболочка слабо выражена, диаметр 3-5 мкм, скорость проведения нервного импульса – 14-25 м/сек (нервы вегетативной нервной системы)
-
Волокна типа С – миелиновой оболочки не имеют, диаметр до 3 мкм, скорость проведения нервного импульса – 2 – 4 м/сек. К ним относятся афферентные волокна, отвечающие за болевую, температурную чувствительность.
Мякотные или безмякотные нервные волокна, объединяясь в пучки, составляют нервный ствол или нерв. Одни из нервов афферентные, другие – эфферентные, но большинство – смешанные, включают и те и другие.
Из безмякотных волокон от 10 до 20% принадлежит симпатическим. Безмякотные преобладают у позвоночных.
Свойства нервных волокон:
-
Возбудимость (у мякотных волокон выше). Хронаксия – от 0,05 мс до 0,2 у двигательных волокон, у симпатических – до 5 мс. Лабильность – 500 – 1000 импульсов в секунду.
-
Изолированное проведение возбуждения. Изоляция обеспечивается миелиновой оболочкой. Нерв состоит из множества нервных волокон, но возбуждение распространяется по каждому волокну отдельно, не переходя на соседние. В безмякотных волокнах возбуждение передается медленно.
-
Двустороннее проведение возбуждения. Импульсы распространяются в обе стороны с одинаковой скоростью.
-
Функционирование только при наличии анатомической и физиологической целостности (при повреждении целостности нервного волокна нарушается изолированное проведение).
-
Неутомимость из-за низкого обмена веществ (Н.Е.Введенский).
-
Скорость проведения возбуждения. В безмякотных нервных волокнах возбуждение распространяется вдоль всей мембраны непрерывно. В мякотных нервных волокнах возбуждение распространяется скачкообразно из-за перехватов Ранвье. Возбуждение проводится при помощи круговых токов. Скорость проведения: мякотные волокна типа А проводят возбуждение со скоростью от 5 до 120 м/с. В мякотных волокна типа В скорость проведения возбуждения – от 3 до 18 м/с, в волокнах типа С – от 0,5 до 3 м/с.
Медиаторы и физиология синапсов. В 1897 году Ч.Шеррингтон ввел понятие «синапс» (от греч. «смыкать», «соединять») для обозначения структуры, обеспечивающей передачу возбуждения с нервного волокна на мышечное или с одного нервного волокна на другое. Это место функционального контакта.
Различают периферические и центральные синапсы.
Периферические находятся на границе между нервными волокнами и рабочими органами.
Центральные – между нервными клетками и их структурами. Обычно встречаются синапсы между концевыми веточками аксона одного нейрона и дендритами (аксо-дендритные синапсы) или телом (аксоматические синапсы) другого нейрона.
В зависимости от природы проходящих по синапсам сигналов различают два типа синапсов – электрические и химические.
Химические синапсы – наиболее распространенный тип синапса у позвоночных. Типичный пример – нервно-мышечный синапс, который существует между окончаниями двигательного нейрона и поверхностью мышечного волокна, это соединение, сходное с межнейронными синапсами в функциональном отношении, но отличающееся по строению. Химический синапс – луковицеобразное утолщение нервных окончаний, названное синаптическими бляшками. Нервно-мышечный синапс состоит из трех основных элементов: пресинаптической мембраны, постсинаптической мембраны и синаптического пространства (Рис.8).
Пресинаптической мембраной служит мембрана нервных веточек – окончаний аксона. Постсинаптическая мембрана – мембрана мышечного волокна. Между ними – синаптическое пространство (щель) шириной примерно 50 нм (рис ).

Рис. Синапс.
1 – везикуля; 2 – митохондрии; 3 - пресинаптическая мембрана; 4 – синаптическая щель; 5 - ионный канал; 6 - постсинапическая мембрана.
Как только в нервное окончание проходит нервный импульс, в синаптических пузырьках начинает выделяться медиатор ацетилхолин, пузырьки подходят к мембране, медиатор выходит из них и попадает в синаптическую щель (в каждом пузырьке около 3000 молекул ацетилхолина). Медиатор – вещество, с помощью которого нервный сигнал передается через синапс. Ацетилхолин – аммонийное производное, выделенное в 1920 г. Отто Леви из окончаний парасимпатических нейронов блуждающего нерва сердца лягушки. На постсинаптической мембране находится особый белок, образующийся в теле клетки на рибосомах - рецептор (холинорецептор), который чувствителен к ацетилхолину. Холинорецептор соединяется с ацетилхолином, структура постсинаптической мембраны изменяется, повышается ее проницаемость для ионов натрия. Проникая из щели в мышечное волокно, ионы натрия вызывают деполяризацию постсинаптической мембраны, возникает потенциал концевой пластинки (только в нервно-мышечном синапсе). При достижении деполяризации определенного уровня (около 40 мВ), между деполяризованным участком постсинаптической мембраны и соседними внесинаптическими участками с прежним потенциалом, возникает движение кругового электрического тока. Если его сила пороговая – возникает распространяющийся потенциал действия (рис. ).

Рис. Последовательность событий при возбуждении синапса.
Ацетилхолинэстераза расщепляет ацетилхолин, в результате гидролиза образуется холин, он всасывается обратно, в синаптическую бляшку, превращается в ацетилхолин, который хранится в пузырьках. Исходное состояние синапса восстанавливается.
Существуют определенные различия в строении и функционировании возбуждающего и тормозного синапсов:
1. Синаптическая щель тормозного синапса уже, она составляет 20 нм (у возбуждающего - 30 нм).
2. Тормозные синапсы имеют более толстую и плотную постсинаптическую мембрану.
3. Пресинаптическая мембрана тормозного синапса содержит меньше пузырьков медиатора.
4. Медиатором в тормозных синапса являются глицин, гамма - аминомасляная кислота (ГАМК), иногда - ацетилхолин.
Для включения в работу тормозного синапса опять нужен импульс возбуждения. При достижении импульсом пресинаптической мембраны в синаптическую щель выбрасывается квант тормозного медиатора. Действуя как химический раздражитель на мембранные рецепторы, тормозной медиатор изменяет проницаемость мембраны незначительно (в основном для ионов K+ и Cl-). Поскольку ионов K+ больше внутри клетки, а ионов Cl- в межклеточном пространстве, они начнут двигаться в противоположных направлениях: K+ - наружу, Cl‾ - внутрь. В результате этого увеличивается поляризованность мембраны (мембранный потенциал). Его продолжительность составляет всего несколько миллисекунд. Но в течение этого времени никакое возбуждение на данном конкретном участке постсинаптической мембраны невозможно. В этом и заключается механизм действия тормозного синапса.
Мозг пользуется несколькими медиаторами. Норадреналин, дофамин, серотонин вызывают возбуждение, глицин и гамма - аминомасляная кислота (ГАМК) - торможение. Ацетилхолин - универсальный медиатор. Он может вызывать возбуждение и торможение. Каждый синапс для своей работы использует только один медиатор.
Электрических синапсов в ЦНС высокоорганизованных животных незначительна. Пресинаптическая мембрана в таких синапсах сообщается с постсинаптической посредством миниатюрных мостиков, т.е. мембраны не разделены щелью. Потенциал действия, достигнув пресинаптической мембраны, преобразуется в градуальный ток, «стекающий» на постсинаптическую мембрану, где вновь возрождается в форме новой волны возбуждения. Таким путем возбуждение проводится через синапс почти без задержки.
Особенностью электрических синапсов является также их способность передавать возбуждение в двух противоположных направлениях, что принципиально невозможно в химических синапсах.
В мозговой ткани встречаются и смешанные синапсы. Такой синапс частично может проводить возбуждение электрическим путем, а частично - с помощью медиатора.
Полагают, что некоторые лекарственные вещества, используемые при депрессиях и тревожности, воздействуют на химическую передачу в синапсах.
Многие транквилизаторы и седативные средства (имипрамин, резерпин, др.) производят лечебный эффект, взаимодействуя с медиаторами, их рецепторами или отдельными ферментами. Галлюциногены типа диэтиламида лизерговой кислоты и москалина воспроизводят действие каких-то природных медиаторов мозга или подавляют действие других медиаторов. Сходный эффект с героином и морфием вызывают эндорфины, способные взаимодействовать с опиатными рецепторами.
Особенности проведения импульса в синапсе:
1. Однонаправленность передачи.
2. Усиление - каждый нервный импульс вызывает освобождение в нервно-мышечном синапсе достаточное количество ацетилхолина, чтобы вызвать распространяющийся ответ в мышечном волокне.
3. Адаптация или аккомодация – при непрерывной стимуляции запасы медиатора будут истощаться, возникает утомление синапса, а значит и торможение передачи сигналов. Адаптивное значение – предотвращается повреждение вследствие перевозбуждения.
Интеграция – способность постсинаптического нейрона суммировать сигналы от пресинаптических нейронов.
-
Дискриминация – временнáя суммация в синапсе позволяет отфильтровать слабые фоновые импульсы, прежде чем они достигнут мозга. Например, экстерорецепторы кожи, глаз и ушей постоянно получают из окружающей сред сигналы, не имеющие особого значения для нервной системы, для нее важны лишь изменения интенсивности стимулов, приводящие к изменению частоты импульсов.
-
Торможение.
Интегрирующая роль центральной нервной системы. Основным механизмом поддержания жизнедеятельности организма на относительно постоянном уровне (гомеостаз) является саморегуляция физиологических функций, осуществляющаяся на основе единства гуморальных и нервных механизмов регуляции.
Гуморальное воздействие осуществляется посредством поступления гормонов и продуктов метаболизма в кровь, лимфу, тканевую жидкость. Нервная система обеспечивает быструю передачу возбуждения и обратную связь между всеми функциональными элементами организма, обеспечивает их соподчинение.
Т.о. осуществляется ее интегративная функция, осуществление которой обеспечивают различные уровни организации ЦНС.
Первый уровень интеграции – нейрон.
Первый уровень интеграции – нейрон, являющийся элементарной единицей ЦНС нейрон, на клеточной мембране которого, как уже отмечалось, и осуществляется интеграция синаптических влияний.
Этот
первый уровень интеграции, характеризующийся
взаимодействием возбуждающих и тормозных
потенциалов, возникает при активации
тормозных входов нейрона.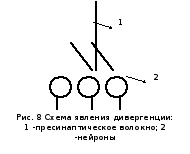
Если возбуждающие и тормозные входы активируются одновременно, происходит суммация синаптических потенциалов противоположной полярности, возможная деполяризация, при которой в клетке (в низкопороговой зоне) возникает потенциал действия.
Второй уровень интеграции – нейронные сети.
Второй уровень интеграции обеспечивается координационной деятельностью элементарных нервных сетей, которая возможна из-за специфики морфологических отношений в ЦНС. А именно: довольно часто наблюдается дивергенция, т.е. явление, когда одно пресинаптическое волокно многократно делится, образуя синаптические контакты сразу со многими нейронами.
Дивергенция обнаруживается практически во всех отделах нервной системы (спинной мозг, афферентный вход, головной мозг, вегетативные ганглии).
В рефлекторных дугах принцип дивергенции функционально проявляется иррадиацией возбуждения, когда раздражение одного афферентного волокна может вызвать генерализованную реакцию за счет возбуждения многих вставочных и моторных нейронов.
В структурной организации нервных сетей встречается ситуация, когда на одном нейроне сходится несколько афферентных терминалей из других отделов ЦНС. Это явление принято называть конвергенцией в нейронных связях (рис.).
К одному мотонейрону, например, могут подходить 6000 коллатералей аксонов первичных афферентов, спинальных интеронейронов, нисходящих из коры и стволовой части мозга.
Все эти окончания образуют на мотонейроне возбуждающие и тормозные синапсы и формируют своеобразную «воронку», ссуженная часть которой представляет общий моторный вход.
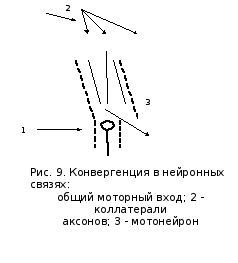
Это анатомическое образование, благодаря которому обеспечивается один из механизмов координационной функции спинного мозга, было открыто английским ученым Ч.Шеррингтоуном, сформулировавшим принцип общего конечного пути. Согласно Шеррингтоуну, количественное преобладание чувствительных и других приходящих волокон над двигательными создает неизбежное столкновение импульсов в общем конечном пути, который представлен группой мотонейронов и иннервируемыми ими мышцами. Из-за столкновения наступает торможение всех степеней свободы двигательного аппарата, кроме той, в направлении которого протекает рефлекторная реакция, вызванная максимальной стимуляцией одного из афферентных входов. Например, одновременно раздражаем рецептивные поля чесательного и сгибательного рефлексов. Импульсы идут от полей к одной и той же группе мотонейронов и здесь, в узком месте воронки, осуществляется выбор в пользу сгибательного рефлекса, вызванного более сильным болевым раздражением. Принцип общего конечного пути действителен для любого этажа ЦНС, даже для моторной коры.
Конвергенция лежит в основе таких физиологических феноменов, как временная и пространственная суммация.
Временнàя суммация – в том случае, если через один афферентный вход к нейрону приходят 2 подпороговых раздражителя. Они следуют один за другим с малым временным интервалом, наступает суммация раздражителей, развивается пороговый уровень, достаточный для генерации импульсной активности.
Биологическое значение этого процесса в том, что он способствует усилению поступающих к нейрону слабых сигналов (и определяется как временная суммация).
В случае если подпороговые раздражители приходят к нейрону через два отдельных входа, возникает суммация ВПСП (возбуждающих) потенциалов, появившихся в двух пространственно разделенных зонах клеточной мембраны. Это явление называется пространственной суммацией (рис. 10)

Рис 10. Схема пространственной суммации:
1 – нейрон; 2 –подпороговые раздражители
1

И временная и пространственная суммация могут вызвать длительную деполяризацию клеточной мембраны.
При
одновременной стимуляции двух входов
нейронов возможен еще один физиологический
феномен – окклюзия.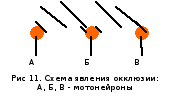
Мотонейрон Б, как видно из схемы, будет возбуждаться дважды – сначала с нейроном А, а затем с нейроном Б. При одновременной стимуляции двух входов нейрон Б возбуждается один раз и соответственно рефлекторный ответ будет меньше алгебраической суммы ответов при раздельной стимуляции. Т.о. феномен окклюзии объясняется наличием дополнительного общего пути для двух входов.
Координирующая функция локальных нервных сетей помимо усиления может проявляться ослаблением слишком интенсивной активности нейронов за счет их торможения.
3-й уровень интеграции.При объединении нескольких локальных сетей формируются нервные центры – комплекс элементов, необходимых для осуществления рефлекса или другого поведенческого акта. Существуют центры дыхания, кровообращения, слюноотделения, глотания, мигания и др.
Нервные центры, даже связанные с одной функцией, могут быть расположены в различных отделах мозга. Например, дыхательный центр представляет собой совокупность нервных образований спинного, продолговатого, среднего, промежуточного мозга и коры больших полушарий.
Эти системы и представляют 3-й интегративный уровень ЦНС.
Деятельность центров базируется на нескольких общих принципах работы.
Принципы работы нервных центров.
Принцип доминанты сформулирован А.А Ухтомским, Под доминантой А.А.Ухтомский понимал господствующий очаг возбуждения, определяющий характер текущих реакций центров в данный момент. Этот господствующий центр может возникнуть при длительном действии гуморальных или рефлекторных раздражителей. Для него характерно: а) наличие повышенной возбудимости; б) иннерционность, обусловленная длительными следовыми процессами; в) способность к суммации и сопряженному торможению других центров, функционально несовместимых с деятельностью центров доминантного очага. Доминанта создаëт определенный вектор поведения.
Примером может служить обнимательный рефлекс у лягушки в результате гормональных воздействий. Гормоны в сочетании с раздражением кожи (рецепторы – мозоли пальцев передних конечностей самца) создают суммацию раздражителей.
А.А.Ухтомский считал, что доминанта является общим принципом работы нервных центров, что она связана с возбуждением или активацией многих нервных центров, временно кооперирующихся для выполнения биологически важной функции. Взаимодействие работающих центров на единый ритм активности (усвоение ритма) рассматривается как динамический функциональный орган.
Согласно современным представлениям (теория функциональных систем П.К.Анохина) любой поведенческий акт, в том числе условный рефлекс – результат анализа и синтеза афферентной информации, включающей, прежде всего, возбуждение центральных рецепторов латерального, переднего гипоталамуса, преоптической области, реагирующих на содержание в крови питательных веществ, половых гормонов и других факторов. Доминирующий очаг может возникнуть не только под влиянием гормональных воздействий, но и под влиянием нервных сигналов, изменяющих возбудимость центральных нейронов.
Иннерционность доминанты - это способность доминантного состояния поддерживаться довольно долго, что обусловлено длительностью следовых процессов. Следовое возбуждение в свете открытий современной физиологии может быть обусловлено следующими факторами:
Суммацией взаимодействия возбуждающих потенциалов, вызванных приходящими к нейронам подпороговыми нервными импульсами.
-
Изменением концентрации ионов К+ в синаптической щели, которые как деполяризаторы, усиливают вхождение ионов Са²+ в пресинаптическое окончание.
-
Следами метаболитов, связанных с медиаторами.
-
Циклическими связями в ЦНС, способными обеспечить следовую самостимуляцию центров (следовые процессы храняться различное время в гиппокампе – часы, сутки – предполагают, что здесь хранятся следовые процессы от пережитых доминат; в спинальных центрах – постсинаптический потенциал длится минуты).
Доминатное состояние как системная реакция может не только суммировать посторонние раздражения, но и может тормозить конкурирующие центры.
Феномен сопряженного торможения конкурирующих центров возможен благодаря динамическому функциональному органу как временной кооперации нервных центров.
Существуют другие теории, объясняющие сопряженное торможение. В 1957 году В.Маунткасл (в опытах использовал кошек, вводя в сенсомоторную кору электроды под углом и перпендикулярно) выдвинул гипотезу колончатой организации коры больших полушарий, согласно которой головной мозг – это совокупность нейронных модулей, связанных многократно. Доминирование одного из модулей и вызывает динамику систем.
Проведение возбуждения через нервные центры осуществляется только в одном направлении: с афферентного нейрона на эфферентный. Свойство объясняется строением синапсов – медиаторы выделяются только концевыми аппаратами аксонов и возбуждение распространяется к постсинаптической мембране.
Задержка проведения в синапсах и время рефлекса обусловлена более медленным проведением нервных импульсов через синапсы, т.к. необходимо время для выделения медиатора, его диффузию через синаптическую щель к постсинаптической мембране и возникновения возбуждающего постсинаптического потенциала под действием медиатора. Чем больше сила раздражителя, тем короче время рефлекса.
Иррадиация возбуждения - это способность возбуждения широкой волной разливаться по ЦНС от центра к центру во всех направлениях, что объясняется наличием огромного количества коллатералей. От каждого аксона отходят идут коллатерали к множеству нейронов, от них идет еще большее число коллатералей. Т.о. импульс, пришедший в ЦНС, может иррадиировать по многим направлениям ко многим центрам.
Конвергенции - свойство нервных центров, противоположное иррадиации. Схождение, сужение – к эфферентному нейрону возбуждение проходит по многим путям, т.к. афферентных путей в 4-5 раз больше.
Циркуляция нервных импульсов по замкнутым нейронным цепям
может длительно осуществляться в одном нервном центре благодаря тому, что импульсы, возникнув в одном из нейронов, передаются к другим нейронам, а затем по коллатералям их аксонов возвращаются на первый нейрон.
Утомляемость нервных центров обуславливается высоким обменом веществ, что приводит к нарушением проведения возбуждения в межнейронных синапсах. Уменьшаются запасы медиатора в окончаниях аксона, падает чувствительность к медиатору постсинаптической мембраны, понижаются ресурсы нервной клетки.
Пластичность нервных центров - перемена функций нервного центра в случае, если рабочий орган, с которым данный орган связан, заменить другим.
Торможение как координирующая функция локальных нервных сетей.
Торможение – это особый нервный процесс, проявляющийся отсутствием способности к активному распространению по нервной клетке (если бы распространению возбуждения с нейрона на нейрон ничего бы не препятствовало, был бы хаос, беспорядочно сокращались бы мышцы, железы).
Различают первичное и вторичное торможение.
Первичное развивается без предварительного воздействия и обусловлено наличием специфических тормозных структур.
Примером может служить возвратное торможение, оно возможно благодаря такому специфическому элементу как клетки Рэншоу. Это вставочный тормозной нейрон в нейронной цепи, в который входит также мотонейрон (рис.12).
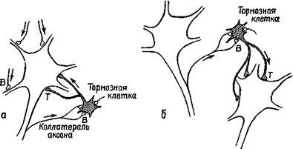
Рис.12. Схема явления возвратного торможения по Рэншоу: 1 – афферентные терминали; 2- тормозной интернейрон; 3 - мотонейрон
Импульсы от возбужденного мотонейрона через отходящие от него возвратные коллатерали также активизируют клетки Рэншоу. А они, в свою очередь, тормозят разряды мотонейроона за счет функции тормозных синапсов, которые клетки Рэншоу образуют на теле активизирующего её нейрона. Т.о. формируется контур с отрицательной обратной связью, позволяющий стабилизировать частоту разрядов моторной клетки и подавать идущую к мышцам избыточную импульсацию. Клетки Рэншоу могут формировать тормозные синапсы не только на активирующих их мотонейронах, но и на соседних со сходными функциями. Такое торможения окружающих клеток называется латеральным.
Рис. Клетки Рэншоу могут формировать тормозные синапсы не только на активирующих их мотонейронах, но и на соседних со сходными функциями. Такое торможения окружающих клеток называется латеральным.
Вторичное торможение – возникает без участия специализированных тормозных структур, как следствие избыточное активизации возбуждающих входов нейрона. Природа торможения – продолжающее действие раздражающего фактора создает в месте воздействия стойкий очаг стойкого стационарного возбуждения, который теряет способность распространяться. Каждое новое раздражение только углубляет этот очаг застойного возбуждения или торможения. Чем сильнее и чаще раздражение, тем глубже торможение, тем сильнее снижается лабильность.