

Тема 8. Функциональная роль коры больших полушарий.
Морфофункциональная организация коры больших полушариев. Кора большого мозга имеет следующие морфофункциональные особенности:
-
многослойность расположения нейронов;
-
модульный принцип организации;
-
соматотопическая локализация рецептирующих систем;
-
экранность, т. е. распределение внешней рецепции на плоскости нейронального поля коркового конца анализатора;
-
зависимость уровня активности от влияния подкорковых структур и ретикулярной формации;
-
наличие представительства всех функций нижележащих структур ЦНС,
-
цитоархитектоническое распределение на поля;
-
наличие в специфических проекционных сенсорных и моторных системах вторичных и третичных полей с ассоциативными функциями;
-
наличие специализированных ассоциативных областей;
-
динамическая локализация функций, компенсация функций утраченных структур;
-
перекрытие в коре большого мозга зон соседних периферических рецептивных полей;
-
возможность длительного сохранения следов раздражения;
-
реципрокная функциональная взаимосвязь возбудительных и тормозных состояний;
-
способность к иррадиации возбуждения и торможения;
-
наличие специфической электрической активности.
Функции отдельных зон новой коры определяются особенностями ее структурно-функциональной организации, связями с другими структурами мозга, участием в восприятии, хранении и воспроизведении информации при организации и реализации поведения, регуляции функций сенсорных систем, внутренних органов. Особенности структурно-функциональной организации коры большого мозга обусловлены тем, что в эволюции происходила кортикализация функций, т. е. передача коре большого мозга функций нижележащих структур мозга.
Однако это не означает, что кора берет на себя выполнение функций других структур. Ее роль сводится к коррекции возможных нарушений функций взаимодействующих с ней систем, более совершенного, с учетом индивидуального опыта, анализа сигналов и организации оптимальной реакции на эти сигналы, формирование в своих и в других заинтересованных структурах мозга памятных следов о сигнале, его характеристиках, значении и характере реакции на него.
Образующие кору нейроны по своему строению и функции подразделяются на несколько групп. Одну группу составляют варьирующие по размеру пирамидные клетки. Они ориентированы вертикально по отношению к поверхности коры и имеют тело треугольной формы. От тела пирамидной клетки вверх отходит длинный Т-образно ветвящийся апикальный дендрит, а вниз от основания нейрона - аксон, который либо покидает кору в составе нисходящих путей, либо направляется к другим зонам коры.
Другая группа корковых нейронов представлена более мелкими звездчатыми клетками. Эти клетки имеют короткие сильно ветвящиеся дендриты и аксоны, формирующие внутрикорковые связи.
Третья группа корковых нейронов включает в себя веретеновидные клетки, имеющие длинный аксон, который ориентирован в горизонтальном или вертикальном направлении.
В связи с тем, что тела и отростки описанных выше нейронов имеют упорядоченное расположение, кора построена по экранному принципу и состоит из шести горизонтальных слоев (рис.)
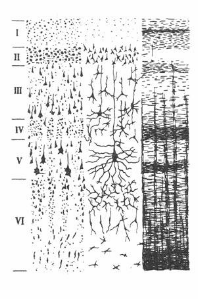
Рис. Строение коры большого мозга.
I. молекулярный, II. наружный зернистый, III. наружный пирамидный,IV. внутренний зернистый, V. ганглиозный (гигантских пирамид), VI. полиморфный.
Самый наружный молекулярный слой слагается из густого сплетения нервных волокон, лежащих параллельно поверхности корковых извилин. Основную массу этих волокон составляют ветвящиеся дендриты пирамидных клеток нижележащих слоев. Сюда же в наружный слой приходят афферентные волокна от таламуса, регулирующих уровень возбудимости корковых нейронов.
Второй слой - наружный зернистый состоит из большого количества мелких звездчатых клеток, которые дополняются малыми пирамидными клетками.
Третий слой - наружный пирамидный - формируется из пирамидных клеток средней величины. Функционально второй и третий слои коры объединяют нейроны, отростки которых обеспечивают кортико-кортикальные ассоциативные связи.
Четвертый слой - внутренний зернистый содержит множество звездчатых клеток. В этом слое преимущественно оканчиваются афферентные волокна, идущие от ядер таламуса.
Пятый слой - внутренний пирамидный образован крупными пирамидными клегками. Наиболее крупные пирамидные нейроны - гигантские клеткш Беца встречаются в прецентральной извилине, занятой моторной зоной коры больших полушарий. Аксоны этих эфферентных корковых нейронов формируют кортикоспинальный (пирамидный) и кортикобульбарные тракты, участвующие в координации целенаправленных двигательных актов и позы.
Шестой – полиморфный слой переходит непосредственно в белое вещество больших полушарий. Этот слой содержит тела нейронов, чьи отростки формируют ксортикоталамические пути.
Наиболее крупные подразделения территории коры — древняя (палеокортекс), старая (архикортекс), новая (неокортекс) и межуточная кора. Поверхность новой коры у человека занимает 95,6%, старой 2,2%, древней 0,6%, межуточной 1,6%.
Если представить себе кору мозга в виде единого покрова (плаща), одевающего поверхность полушарий, то основная центральная часть его составит новая кора, в то время как древняя, старая и межуточная займут место на периферии, т. е. по краям этого плаща.
Древняя кора состоит из одного клеточного слоя, нечетко отделённого от нижележащих подкорковых ядер. Старая кора полностью отделена от последних и представлена 2—3 слоями. Новая кора состоит, как правило, из 6—7 слоев клеток. Межуточные кора — переходные структуры между полями старой и новой коры, а также древней и новой коры — из 4—5 слоев клеток.
Основной тип прямых и обратных связей новой коры — вертикальные пучки волокон, приносящие информацию из подкорковых структур к коре и посылающие её от коры в эти же подкорковые образования.
Наряду с вертикальными связями имеются внутрикортикальные — горизонтальные пучки ассоциативных волокон, проходящие на различных уровнях коры и в белом веществе под корой.
Горизонтальные пучки наиболее характерны для I и III слоев коры, а в некоторых полях для V слоя. Горизонтальные пучки обеспечивают обмен информацией как между полями, расположенными на соседних извилинах, так и между отдалёнными участками коры (например, лобной и затылочной).
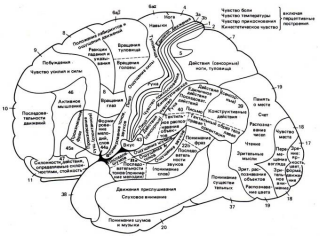
Рис. 6.1. Разделение латеральной коры головного мозга на функциональные поля по Клайсту. Цифрами обозначены цитоархитектонические поля Бродмана.
Выраженность отдельных слоев в различных областях коры не одинакова. Учитывая эту особенность, К. Бродман по гистологическим признакам, в частности по плотности расположения и форме нейронов, разделил всю кору на 50 цитоархитектонических полей (рис..).
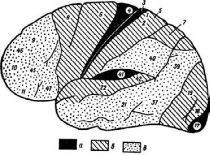
Рис. 4. Карта цитоархитектонических полей коры головного мозга.
Конвекситальная поверхность коры больших полушарий: а — первичные поля; б — вторичные поля; в — третичные поля
Позднее были разработаны функциональные принципы классификации различных зон коры. При этом оказалось, что зоны, выделенные на основании их функциональных и нейрохимических особенностей, в известной степени соответствуют цитоархитектоническому разделению коры на поля.
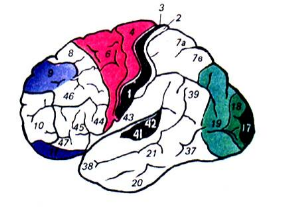
Рис.. Цитоархитектонические поля коры больших полушарий человека.
Так, например, при сравнении наиболее изученных сенсорных и моторных зон коры оказалось, что в первых наружный пирамидный слой (3) выражен слабо и доминируют зернистые слои (2, 4), где оканчиваются сенсорные афференты (гранулярная кора). И, напротив, в моторных зонах коры зернистые слои развиты плохо (агранулярная кора), а пирамидные слои превалируют.
Таким образом, функциональная специализация накладывает определенный отпечаток на структуру сенсорных и моторных зон коры, и выделение этих областей по различным системам классификации не случайно.
Проекционные зоны коры. Использование методов удаления и раздражения отдельных зон коры позволило идентифицировать ее моторные (мотосенсорные) области. Важнейшая двигательная область коры локализована в прецентральнои извилине (поля 4 и 6, по Бродману), лежащей кпереди от центральной (роландовой) борозды, которая отделяет лобную долю от теменной (рис.15.1.).
В 30-х гг. нашего столетия У. Пенфилд, установил наличие правильной пространственной проекции соматических мышц различных отделов тела на двигательную область коры. Обнаружив соматотопическую организацию двигательной коры рис. ). Относительные размеры органов отражают ту площадь коры головного мозга, с которой могут быть вызваны соответствующие ощущения и движения. У. Пенфилд отметил, что зоны, управляющие движениями кистей рук и мимической мускулатурой лица, являются наиболее обширными.
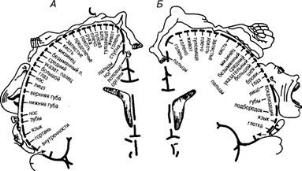
Рис. 11. Схема соматотопической проекции общей чувствительности и двигательных функций в коре головного мозга (по У. Пенфилду):
А — корковая проекция общей чувствительности; Б — корковая проекция двигательной системы.
В дальнейшем на медиальной поверхности полушария рядом с этой первичной моторной областью была обнаружена еще одна вторичная двигательная область и, таким образом, было показано множественное представительство периферических образований в нескольких двигательных зонах коры.
В связи с тем, что эти области помимо моторного выхода из коры имеют самостоятельные сенсорные входы от кожных и мышечных рецепторов, они были названы первичной, к вторичной мотосенсорной корой.
В лежащей позади роландовой борозды постцентральной извилине (поля 1, 2, 3) находится первая соматосенсорная (сенсомоторная) область, куда приходят афферентные проекции от рецепторов кожи и двигательного аппарата (рис. ). Так же как и в двигательной коре, эти проекции имеют соматотопическую организацию, причем близкие друг от друга точки на поверхности тела имеют представительство в соседних участках постцентральной извилины.
Кроме первой соматосенсорной области обнаружена вторая соматосенсорная область, локализованная вентральнее в районе сильвиевой борозды, которая отделяет височную долю от теменной и лобной. Здесь также имеется соматотопическая проекция поверхности тела, однако менее четкая.
Оказалось, что первая и вторая соматосенсорные области кроме афферентных входов содержат моторные выходы и, следовательно, их правильнее называть первичной и вторичной сенсомоторными зонами (рис.).
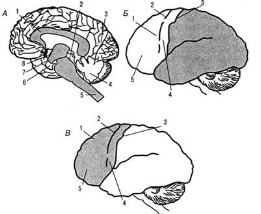
Рис. 9. Структурно-функциональная модель интегративной работы мозга (по А. Р.Лурия, 1970):
А) I блок — регуляции общей и избирательной неспецифической активации мозга, включающий ретикулярные структуры ствола, среднего мозга и диэнцефальных отделов, а также лимбическую систему и медиобазальные
отделы коры лобных и височных долей мозга: 1 — мозолистое тело, 2 — средний мозг, 3 — теменно-затылочная борозда, 4 — мозжечок, 5 — ретикулярная формация ствола, 6 — крючок, 7 — гипоталамус, 8 — таламус;
Б) II блок — приема, переработки и хранения экстероцептивной информации, включающий основные анализаторные системы (зрительную, кожно-кинестетическую, слуховую), корковые зоны которых расположены в задних отделах больших полушарий: 1 — премоторная область, 2 — прецентральная извилина, 3 — центральная извилина, 4 — моторная область, 5 — префронтальная область;
В) III блок — программирования, регуляции и контроля за протеканием психической деятельности, — включающий моторные, премоторные и префронтальные отделы мозга с их двухсторонними связями.
Следовательно, можно судить о наличии в коре четырех чувствительных областей, расположенных в порядке убывания их значимости.
Равным образом в коре существует четыре двигательных области, причем ведущая роль в этом ряду принадлежит первичной мотосенсорной коре.
Из числа других проекционных зон коры следует отметить первичную зрительную область (поле 17), локализованную в затылочной доле. В проекциях рецепторов сетчатки на поле 17, так же как и в других сенсорных системах, обнаруживается точный топографический порядок, причем в каждое полушарие проецируются одноименные половины сетчаток (левые половины - в правое полушарие, а правые - в левое). Происходящее при этом в каждом полушарии совмещение зрительных полей обоих глаз лежит в основе бинокулярного зрения.
Рядом с первичной зрительной областью расположена вторичная, занимающая поля 18 и 19. Эти поля имеют отношение к таким функциям, как зрительное внимание и управление движениями глаз .
В настоящее время хорошо изучена первичная проекционная зона слухового анализатора, которая локализована в верхнем крае височной доли (поля 41 и 42). Для этой области, так же как и для других проекционных зон, описаны участки представительства кортиева органа улитки, состоящее в том, что различные участки кортиева органа проецируются в определенные локусы слуховой коры.
В пределах каждой доли коры больших полушарий рядом с проекционными зонами расположены поля, которые не связаны с выполнением какой-либо специфической сенсорной или моторной функции. Такие поля составляют ассоциативную кору, для нейронов которой свойственно отвечать на раздражение различных модальностей и таким образом участвовать в интеграции сенсорной информации и в обеспечении связей между чувствительными и двигательными зонами коры. Эти механизмы являются физиологической основой высших психических функций.
В состав ассоциативный коры входит ряд областей теменной, височной и лобной долей. К сожалению, границы этих областей и кортико-кортикальные связи между ними идентифицированы не полностью, а большинство физиологических данных базируется на симптомах поражения ассоциативных областей у животных и на данных клинических наблюдений.
Вместе с тем в настоящее время установлено, что теменные ассоциативные поля, которые в процессе эволюции надстраиваются над зрительной проекционной зоной, участвуют в оценке биологически значимой информации и в восприятии пространственных отношений окружающего мира рис. ).
Лобные доли (поля 9-14), имеющие обширные двусторонние связи с лимбической системой мозга, контролируют оценку мотивации поведения и программирование сложных поведенческих актов. Установлено участие лобных долей в управлении движениями.
Колончатая организация зон коры. Применение современных микроэлектродных методов в значительной мере расширило представления о переработке сенсорной информации в неокортексе.
Соматосенсорная кора организована в элементарные функциональные единицы - колонки, ориентированные перпендикулярно поверхности. Колонка является элементарным блоком сенсомоторной коры, где осуществляется локальная переработка информации от рецепторов одной модальности.
Согласно современным представлениям, каждая функциональная колонка сенсомоторной коры состоит из нескольких морфологических микромодулей, объединяющих пять-шесть нейронов.
На рис. приводится возможная схема микромодуля сенсомоторной коры. В его состав входит несколько пирамидных клеток, дендриты которых максимально сближены и образуют дендритный пучок; в пределах этого пучка возможны электротонические связи, которые обеспечивают, по всей вероятности, синхронную работу всего объединения.
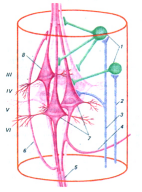
Рис.15.2. Структурный модуль сенсомоторной коры:
III - YI -корковые слои; 1 - интернейроны, 2 - афферентный вход, контактирующий с интернейроном, 3 - афферентный вход, образующий терминали на пирамидных нейронах, 4 - возвратная коллатераль аксона, вступающая в контакт с тормозным интернейроном, 5 - пучок аксонов, выходящий за пределы колонки, 6 - возвратная коллатераль, обеспечивающая облегчающие влияния в пределах модуля, 7 - кортикоспинальные пирамидные клетки, 8 - кортикорубральная пирамидная клетка.
К группе ориентированных по вертикали пирамидных клеток примыкают звездчатые клетки, с которыми контактируют приходящие к микромодулю таламокортикальные волокна. Некоторые из звездчатых клеток, выполняющие тормозную функцию, имеют длинные аксоны, распространяющиеся в горизонтальном направлении.
Аксоны пирамидных клеток образуют возвратные коллатерали, которые могут обеспечивать и облегчающие влияния в пределах микромодуля, и тормозные взаимодействия между микромодулями, контактируя с тормозными интернейронами.
Несколько структурных микромодулей, объединенных горизонтальным ветвлением терминалей специфических таламокортикальных афферентов, аксоны терминалей и отростков звездчатых клеток формируют колонку (или макромодуль). Каждая колонка характеризуется функциональным единством, которое проявляется в том, что нейроны колонки отвечают на раздражитель одной модальности.
В пределах одной колонки нейроны можно дифференцировать по характеру их импульсных ответов на простые и сложные.
У простых нейронов распределение импульсов в ответе на раздражение адекватного механорецептора идентично ответу самого рецептора.
Сложные нейроны реагируют на периферические стимулы иначе, чем связанные с ними рецепторы, и дают максимальный ответ только на определенные формы раздражения, например на движущиеся стимулы. Сложные нейроны связаны с более поздней ступенью анализа тактильного восприятия.
Нейрофизиологические исследования сенсомоторной коры показали, что обычно возбуждение одного из модулей сопровождается торможением соседних. Очевидно, этот факт свидетельствует о том, что процесс торможения препятствует иррадиации возбуждения на смежные модули и, таким образом, усиливает контрастное различение контуров тактильного раздражения.
Изучение других проекционных зон коры обнаружило колончатою организацию зрительной коры. В зрительной коре наблюдается регулярное чередование колонок, нейроны которых реагируют на оптическое раздражение либо только правого, либо только левого глаза. Кроме этих глазодоминантных колонок в зрительной коре обнаружен другой набор колонок, отличающихся друг от друга осевой ориентацией рецептивного поля.
Колончатый принцип организации нейронов присущ и слуховой коре, где колонки дифференцируются по способности суммировать информацию, идущую от двух ушей.
Наконец, в мотосенсорной коре также были обнаружены колонки, способные возбуждать или тормозить группу функционально однородных мотонейронов.
Таким образом, колончатая организация является общей чертой структуры мотосенсорных и сенсомоторных зон коры.
Возможно, в первичных проекционных областях этот принцип организации интимно связан с процессами, создающими в мозгу отображение различных параметров внешнего раздражителя. Однако вопросы о том, как используется эта информация за пределами проекционных зон, как из информации об отдельных качествах внешнего объекта мозг синтезирует цельную картину реальности, пока остаются открытыми.
По всей вероятности, в мозгу должна существовать система, которая считывает элементарные процессы в колонках и объединяет дробную информацию в более сложные сочетания. Реальных данных о существовании такой системы нейронов нет, однако некоторые гипотезы о возможных механизмах синтетической деятельност мозга существуют.
Одна из них принадлежит Дж. Экклсу. Согласно этой гипотезе, каждая корковая колонка, или модуль, может попеременно участвовать в различных сложных системах модулей.
Формирование такой сложной системы обусловлено внутрикортикальными связями между отдельными модулями, которые могут иметь и возбуждающий, и тормозной характер. Активирующие воздействия на модуль в первую очередь могут поступать через: поверхностные горизонтальные волокна, которые образуют синапсы на апикальных дендритах крупных пирамидных клеток.
Эта активация может усиливаться за счет импульсации, идущей в модуль по таламокортикальным входам. В то же время через тормозные интернейроны активность соседних модулей может быть подавлена.
Таким образом, баланс возбуждающих и тормозных процессов в данный момент может выступать в роли координирующего фактора, который определяет формирование сложных систем модулей.
Согласно современным представлениям, такая система расположенных в разных отделах мозга и связанных единством функций модулей носит название распределенной.
С точки зрения отмеченных выше соображений, сегодня старое представление о локализации функций не выдерживает критики. Даже такой простой поведенческий акт, как захватывание предмета пальцами, предполагает содружественную работу центров зрительной, моторной коры и лобной доли, которая участвует в регуляции глазных движений. Вполне естественно, что управление высшими психическими функциями тем более нельзя считать компетенцией какого-то одного центра. Реализацию каждой функции осуществляет распределенная система, состоящая из корковых и подкорковых центров, соединенных между собой нервными связями.
Базальные (подкорковые) ядра головного мозга располагаются под белым веществом внутри переднего мозга, преимущественно в лобных долях. К базальным ядрам относят хвостатое ядро, скорлупу, ограду, бледный шар (рис. ).
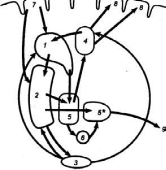
Рис. 3.29 Афферентные и эфферентные связи базальных ядер 1 — хвостатое ядро, 2 — скорлупа, 3 — черное вещество, 4 — таламус, 5, 5* - внутренний и внешний отделы бледного шара, 6 - субталамическое ядро, 7 — сенсомоторная кора, 8 - двигательная кора, 9 - пути к стволовой части мозга; стрелками обозначены афферентные пути, эфферентные пути, межъядерные взаимодействия.
Хвостатое ядро и скорлупа являются эволюционно более поздними, чем бледный шар, образованиями и функционально оказывают на него тормозящее влияние.
Функции любых образований головного мозга определяются, прежде всего, их связями, которых у базальных ядер достаточно много. Эти связи имеют четкую направленность и функциональную очерченность.
Хвостатое ядро и скорлупа получают нисходящие связи преимущественно от экстрапирамидной коры. Другие поля коры большого мозга также посылают большое количество аксонов к хвостатому ядру и скорлупе.
Основная часть аксонов хвостатого ядра и скорлупы идет к бледному шару, отсюда - к таламусу и только от него - к сенсорным полям.
Следовательно, между этими образованиями имеется замкнутый круг связей.
Хвостатое ядро и скорлупа имеют также функциональные связи со структурами, лежащими вне этого круга: с черной субстанцией, красным ядром, мозжечком, спинным мозгом и др.
Обилие и характер связей хвостатого ядра и скорлупы свидетельствуют об их участии в интегративных процессах, организации и регуляции движений, регуляции работы вегетативных органов.
Хвостатое ядро и бледный шар принимают участие в таких интегративных процессах, как условно-рефлекторная деятельность, двигательная активность. Прямое раздражение некоторых зон хвостатого ядра вызывает поворот головы в сторону, противоположную раздражаемому полушарию.
Раздражение других областей хвостатого ядра и скорлупы вызывает прекращение всех видов активности: ориентировочной, эмоциональной, двигательной, пищевой.
Раздражение хвостатого ядра может полностью предотвратить восприятие болевых, зрительных, слуховых и других видов стимуляции. При стимуляции хвостатого ядра удлиняются латентные периоды рефлексов, нарушается переделка условных рефлексов. Выработка условных рефлексов на фоне стимуляции хвостатого ядра становится невозможной.
Видимо, это объясняется тем, что стимуляция хвостатого ядра вызывает торможение активности коры большого мозга. Ряд подкорковых структур также получает тормозное влияние хвостатого ядра.
Таким образом, специфичным для раздражения хвостатого ядра является преимущественно торможение активности коры большого мозга, подкорковых образований, торможение безусловного и условно-рефлекторного поведения.
В то же время, при раздражении хвостатого ядра могут появляться некоторые виды изолированных движений. Видимо, хвостатое ядро имеет наряду с тормозящими и возбуждающие структуры.
В случае повреждения хвостатого ядра наблюдаются существенные расстройства высшей нервной деятельности, затруднение ориентации в пространстве, нарушение памяти, замедление роста организма.
Несмотря на большое функциональное сходство хвостатого ядра и скорлупы, имеется ряд функций, специфичных последней. Для скорлупы характерно участие в организации пищевого поведения: пищепоиска, пищенаправленности, пищезахвата и пищевладения; ряд трофических нарушений кожи, внутренних органов.
Функция хвостатого ядра не является просто тормозной, а заключается в корреляции и интеграции процессов оперативной памяти. Это подтверждается также тем, что на нейронах хвостатого ядра конвергирует информация различных сенсорных систем, так как большая часть этих нейронов полисенсорна.
Бледный шар связан с таламусом, скорлупой, хвостатым ядром, средним мозгом, гипоталамусом и др., что свидетельствует об его участии в организации простых и сложных форм поведения.
Раздражение бледного шара вызывает сокращение мышц конечностей, активацию или торможение мотонейронов спинного мозга. Стимуляция бледного шара в отличие от стимуляции хвостатого ядра не вызывает торможения, а провоцирует ориентировочную реакцию, движения конечностей, пищевое поведение (обнюхивание, жевание, глотание и т.д.).
Ограда содержи нейроны разных видов. Стимуляция ограды вызывает ориентировочную реакцию, поворот головы в сторону раздражения, жевательные, глотательные, иногда рвотные движения. Раздражение ограды тормозит условный рефлекс на свет. Стимуляция ограды во время еды тормозит процесс поедания пищи.
Известно, что толщина ограды левого полушария у человека несколько больше, чем правого; при повреждении ограды правого полушария наблюдаются расстройства речи.
Таким образом, базальные ядра головного мозга являются интегративными центрами организации моторики, эмоций, высшей нервной деятельности, причем каждая из этих функций может быть усилена или заторможена активацией отдельных образований базальных ядер.