

Тема 6. Функциональное значение различных отделов центральной нервной системы.
Спинной мозг - самый древний и примитивным отделом ЦНС, сохраняющим свою сегментарность.
Первая особенность сегментарной иннервации состоит в том, что каждый сегмент спинного мозга иннервирует 3 метамера (сегмента тела) - собственный, половину вышележащего и половину нижележащего сегмента. Получается, что каждый метамер получает иннервацию от трех сегментов спинного мозга. Это гарантирует выполнение спинным мозгом его функций при повреждении мозга и его корешков.
Вторая особенность сегментарной иннервации заключается в избытке чувствительных волокон в составе задних корешков спинного мозга по сравнению с количеством двигательных волокон передних корешков. При большом разнообразии поступающей информации с периферии организм использует небольшое количество исполнительных структур для ответной реакции.
Афферентные волокна несут импульсы от рецепторных полей кожи и мышц шеи, туловища конечностей; внутренних органов.
Эфферентные волокна несут импульсы от ЦНС к исполнительным органам (мышцам и железам (рис. )
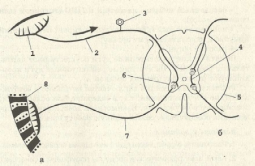
Рис. . Рефлекторная дуга (а — двухнейронная, б — трехнейронная):
1 — рецептор; 2 — чувствительный (центростремительный) нерв; 3 — чувствительный нейрон в спинно-мозговом ганглии; 4 — вставочный нейрон; 5 — спинной мозг; 6 — двигательный нейрон в передних рогах спинного мозга; 7 — двигательный (центробежный) нерв; 8 — рабочий орган
Мотонейроны спинного мозга возбуждаются за счет афферентных импульсов, идущих от рецепторных полей. Активность мотонейронов зависит не только от потоков афферентной информации, но и от сложных внутрицентральных взаимоотношений.
Важную роль играют нисходящие влияния коры полушарий, подкорковых ядер и ретикулярной формации, которые корректируют спинальные рефлекторные реакции.
Большое значение имеют многочисленные контакты вставочных нейронов, среди которых особая роль принадлежит тормозным клеткам. Образуя тормозные синапсы, они контролируют работу мотонейронов и предупреждают их перевозбуждение. В работу нейронов вмешиваются также потоки импульсов обратной афферентации, идущие от проприорецепторов мышц.
Вставочные нейроны спинного мозга за счет богатства синаптических связей обеспечивает собственную интегративную деятельность спинного мозга, включая управление сложными двигательными актами.
Объем функций, осуществляемых спинным мозгом, чрезвычайно велик. В нем находятся центры всех двигательных рефлексов (за исключением мускулатуры головы), всех рефлексов мочеполовой системы и прямой кишки, рефлексов, обеспечивающих терморегуляцию, регулирующих метаболизм тканей, центры большинства сосудистых рефлексов, центр сокращения диафрагмы и др. В естественных условиях эти рефлексы всегда испытывают влияние высших отделов головного мозга.
Рефлекторная функция спинного мозга включает защитные рефлексы, рефлексы на растяжение, мышц-антагонистов, висцеромоторные, вегетативные рефлексы. Эта классификация весьма условна, и весь ее смысл в том, что она указывает на многообразие рефлекторных ответов.
Рефлексы растяжения проявляются укорочением мышцы в ответ на ее растяжение. Основными рецепторами в этом случае служат нервно-мышечные веретена, афферентным звеном - чувствительные волокна соматических нервов и дорсальных корешков спинного мозга. Эти рефлекторные дуги чаще всего замыкаются в спинном мозге.
Биологическое значение этих рефлексов состоит в том, что они участвуют в сохранении статики и положения тела, регулируя степень сокращения мышцы в соответствии с падающими на нее раздражениями.
Рефлексы мышц-антагонистов лежат в основе локомоторных актов и характеризуются тем, что при возбуждении мотонейронов сгибателей одновременно происходит торможение мотонейронов мыщц-разгибателей. При этом в конечности другой стороны наблюдаются обратные явления. В целом это создает правильное чередование противоположных по функциональному значению мышечных сокращений.
Висцеромоторные рефлексы возникают при возбуждении афферентных волокон внутренних органов и характеризуются появлением двигательных реакций мышц грудной и брюшной стенки, мышц-разгибателей спины.
Вегетативные рефлексы проявляются в ответ на возбуждение симпатических и соматических чувствительных клеток и в ответ на раздражение чувствительных путей.
Наряду с выполнением собственных рефлекторных реакций нейронные структуры спинного мозга служат аппаратом для реализации большого числа сложных процессов, осуществляемых различными отделами головного мозга. Это управление может быть прямым, когда нисходящие пути непосредственно связаны с мотонейронами спинного мозга, и непрямым через интернейроны, образующие короткие межсегментарные связи.
Проводниковые функции спинного мозга осуществляются белым веществом, состоящим из нервных волокон.

Рис.9.2. Расположение пучков проводящих путей (на поперечном разрезе спинного мозга)
1 - нежный, 2 - клиновидный, 3, 4 - задний и передний мозжечковые, 5, 6 - латеральный и вентральный спиноталамическне, 7 - спиннотекгальный, 8 - пирамидный, 9 - оливоспинальный, 10 - вестибулоспинальный, 11 - руброспинальный, 12 -перекрещенный пирамидный путь.
Под проводящими путями принято понимать группы нервных волокон, характеризующиеся общностью строения и функций. Они связывают различные отделы спинного мозга или спинной и головной мозг.
Восходящие проводящие пути несут импульсы от рецепторов, воспринимающих информацию из внешней и внутренней среды организма. В зависимости от вида чувствительности, которую они проводят, их делят на пути экстеро-, проприо- и интероцептивной чувствительности.
Нисходящие пути передают импульсы от структур головного мозга к двигательным ядрам, осуществляющим ответные реакции на внешние и внутренние раздражения.
Основными восходящими путями спинного мозга являются тонкий пучок, клиновидный пучок, латеральный и вентральный спиноталамические тракты, дорсальный и вентральный спинно-мозжечковые тракты (рис.).
Тонкий и клиновидный пучки составляют задние столбы спинного мозга. Эти пучки волокон являются отростками чувствительных клеток спинальных ганглиев, проводящих возбуждение от проприоцепторов мышц сухожилий, частично тактильных рецепторов кожи, висцерорецепторов. По пути они отдают большое число ветвей к нейронам вышележащих сегментов спинного мозга, образуя, межсегментарные связи.
Считается, что по этой системе проводится информация тонко дифференцированной чувствительности, позволяющая определить локализацию, контур периферического раздражения, а также его изменения во времени.
По латеральному и вентральному спиноталамическим трактам проводится болевая, температурная и тактильная чувствительность Полагают, что по этой системе передается информация о качественной природе раздражителей.
Дорсальный спинно-мозжечковый тракт - чувствительный путь спинного мозга, идущий к мозжечку. По ним проводится информация от рецепторов мышц и связок конечностей.
Вентральный спинно-мозжечковый тракт направляются к коре мозжечка. Они участвуют в поддержании тонуса мышц для выполнения движений и сохранения позы.
Нисходящие проводящие пути. Эти пути связывают высшие отделы ЦНС с эффекторными нейронами спинного мозга. Основными из них являются пирамидный, руброспинальный, вестибулоспинальный и ретикулоспинальный тракты.
Пирамидный тракт образован аксонами клеток двигательной зоны коры больших полушарий. Направляясь к продолговатому мозгу, эти аксоны отдают большое число коллатералей структурам промежуточного, среднего, продолговатого мозга и ретикулярной формации. В нижней части продолговатого мозга большая часть волокон пирамидного пути переходит на противоположную сторону (перекрест пирамид), образуя латеральный пирамидный тракт. В спинном мозгу он располагается в боковом столбе.
Другая часть волокон идет, не перекрещиваясь, до спинного мозга и только на уровне сегмента, в котором оканчивается, переходит на противоположную сторону. Это прямой пирамидный тракт. Оба они заканчиваются на мотонейронах передних рогов серого вещества спинного мозга.
Основной функцией пирамидных путей является передача импульсов для выполнения произвольных движений.
Вестибулоспинальный тракт образован волокнами, ядра которых лежат в продолговатом мозгу. По нему передаются импульсы, регулирующие тонус мускулатуры, согласованность движений, равновесие.
В спинном мозгу помимо длинных имеются и короткие нисходящие пути, соединяющие между собой его отдельные сегменты.
Продолговатый мозг является непосредственным продолжением вверх спинного мозга. Продолговатый мозг, мост, средний и промежуточный мозг образуют ствол мозга (рис. ).
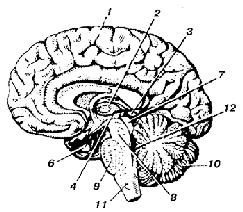
Рис. Головной мозг человека (правая половина, вид слева)
1 - большое полушарие; 2 - зрительный бугор (таламус);3 - надбугорье (эпиталамус); 4 - подбугорье (гипоталамус);5 - мозолистое тело; 6 - гипофиз; 7 - четверохолмие;8 - ножки мозга; 9 - варолиев мост; 10 – мозжечок; 11 - продолговатый мозг;12 - четвёртый желудочек головного мозга.
Задний мозг, состоящий из продолговатого мозга и Варилиева моста, представляет собой отдел головного мозга, который является непосредственным продолжением спинного мозга.
Серое вещество спинного мозга переходит в серое вещество продолговатого мозга и сохраняет черты сегментарного строения. Однако основная часть серого вещества распределена по всему объему заднего мозга в виде обособленных ядер, разделенных белым веществом.
В продолговатом мозгу симметрично заложены ядра четырех последних пар черепно-мозговых нервов, осуществляющих афферентную и эфферентную иннервацию головы и внутренних органов.
В самой нижней части ромбовидной ямки располагается ядро подъязычного нерва (XII пара), иннервирующего мышцы языка. Двигательное ядро добавочного нерва (XI пара) является продолжением заднебокового отдела переднего рога спинного мозга и связано с иннервацией мускулатуры шеи (рис.10.1.).
Группа ядер блуждающего нерва (X пара) участвуют в вегетативной, двигательной и чувствительной иннервации. Вегетативное ядро дает волокна к гортани, пищеводу, сердцу, желудку, тонкой кишке, пищеварительным железам. Чувствительные волокна от многих внутренних органов заканчиваются в чувствительном ядре. Третье ядро блуждающего нерва регулируют последовательность сокращения мышц глотки и гортани при дыхании.
Языкоглоточный нерв (IX пара), так же как и блуждающий, содержит двигательные, чувствительные и вегетативные волокна. Двигательные волокна участвуют в иннервации мышц ротовой полости и глотки. Чувствительные волокна иннервируют вкусовые рецепторы языка, слизистой оболочки глотки. Вегетативное ядро языкоглоточного нерва, связано с иннервацией секреторного аппарата слюнных желез.
Слуховой, или вестибулокохлеарный нерв (VIII пара) состоит из двух частей: нерва улитки и нерва преддверия. Волокна нерва улитки, или собственно слухового нерва, представляют собой аксоны клеток лежащего в улитке спирального ганглия.
Волокна нерва преддверия формируются аксонами клеток узла преддверия, локализованного на дне внутреннего слухового прохода.
Варолиев мост, являющийся структурой заднего мозга, имеет вид поперечно лежащего утолщенного валика.
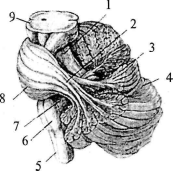
Рис. . Расположение мозжечка на стволе мозга (часть ткани мозжечка убрана):
1 — верхняя ножка мозжечка; 2 — средняя ножка мозжечка; 3 — лист (долька мозжечка); 4 — левое полушарие мозжечка; 5 — спинной мозг; 6 — продолговатый мозг; 7 — нижняя ножка мозжечка; 8 — мост; 9 — средний мозг
Ниже моста находится продолговатый мозг, выше располагается средний мозг.
Вентральная поверхность Варолиева моста представляет собой мощную поперечно-волокнистую выпуклость белого вещества. От Варолиевого моста отходят четыре пары черепно-мозговых нервов: тройничный нерв (V пара), отводящий нерв (VI пара), лицевой нерв (VII пара), предверно-улитковый, или слуховой, нерв (VIII пара).
Передняя часть моста состоит из нервных волокон, образующих нисходящие проводящие пути, среди которых находятся клеточные скопления – ядра. Проводящие пути моста связывают кору большого мозга со спинным мозгом, с двигательными ядрами черепных нервов и с корой полушарий мозжечка.
Дорсальная часть моста является непосредственным продолжением продолговатого мозга. В ней располагаются переключательные ядра сенсорных систем, ядра черепно-мозговых нервов и ретикулярной формации.
Функции Варолиева моста. В латеральном отделе моста расположено ядро лицевого нерва (VII пара). Этот нерв является смешанным. Афферентные чувствительные волокна передают информацию от вкусовых рецепторов языка. Вегетативные волокна участвуют в иннервации подчелюстных и подъязычных слюнных желез. Эфферентные волокна лицевого нерва имеют концевые разветвления в мимической мускулатуре лица.
Ядро отводящего нерва (VI пара) заложено поверхностно в задней половине моста. Моторные волокна этого нерва иннервируют мышцы, двигающие глазное яблоко, чувствительные волокна связаны с проприоцепторами этих мышц (рис.10.1.).
И наконец, последняя пара черепно-мозговых нервов, относящаяся к заднему мозгу, - это тройничный нерв (V пара). Двигательное ядро участвует в иннервации жевательных мышц, мышцы нёбной занавески и мышцы, напрягающей барабанную перепонку. Длинное, простирающееся через весь мост чувствительное ядро тройничного нерва содержит клетки, к которым подходят аксоны от рецепторов кожи лица, слизистой оболочки носа, зубов, надкостницы костей черепа.

Рис.10.1. Расположение ядер черепно-мозговых нервов в стволовой части мозга:
1 - добавочный нерв, 2 - подъязычный, 3 - слуховой, 4 - отводящий, 5 - блоковый, 6 - глазодвигательный, 7 - тройничный, 8 - лицевой, 9 - языкоглоточный, 10 - блуждающий нерв.
В заднем мозге имеются ядра чувствительных путей задних столбов спинного мозга - ядра тонкого и клиновидного пучков, являющихся путями, которые проводят информацию о положении тела в пространстве.
Рефлексы Варолиева моста. При участии ядер заднего мозга осуществляются цепные рефлексы, связанные с жеванием и проглатыванием пищи.
С функцией пищеварительного тракта связаны многие вегетативные рефлексы заднего мозга. К их числу относится, например, рефлекторная регуляция секреции слюнных желез, которая осуществляется с участием ядер лицевого и языкоглоточного нервов, содержащих парасимпатические нейроны. Вегетативное ядро блуждающего нерва включено в систему рефлекторной регуляции дыхания, деятельности сердца и тонуса сосудов.
Целая группа рефлексов заднего мозга направлена на поддержание позного тонуса. В зависимости от источника рецепторных воздействий тонические рефлексы подразделяются на шейные и вестибулярные, или лабиринтные.
Шейные тонические рефлексы запускаются при возбуждении проприоцепторов мышц шеи. При наклоне головы вниз тонус разгибателей передних конечностей снижается, а задних - возрастает. Нарушение равновесия за счет поворота головы вправо или влево относительно продольной оси вызывает компенсаторное усиление тонуса разгибателей тех конечностей, в сторону которых повернута голова.
Вестибулярные рефлексы дополняют шейные тонические рефлексы. Вестибулярные рефлексы не зависят от положения головы относительно туловища.
Статокинетические рефлексы направлены на поддержание позы при изменении скорости движения. Например, ускорение при вращении тела в горизонтальной плоскости возбуждает рецепторы горизонтального полукружного канала и вызывает рефлекторную реакцию глазодвигательного аппарата - горизонтальный нистагм. Суть этой реакции состоит в том, что в момент ускорения вращения глаза движутся в сторону, противоположную направлению вращения. Затем, достигнув крайнего отклонения, глаза быстро перемещаются обратно в направлении вращения и таким образом в поле зрения попадает другой участок пространства.
Ретикулярная (сетевидная) формация занимает центральную часть заднего мозга. Ретикулярные нейроны распределяются диффузно и группируются в ядрах. По своей топографии ретикулярная формация ствола мозга является структурой, расположенной на пути всех входных и выходных систем мозга.
К ретикулярным нейронам подходят коллатерали сенсорных восходящих путей стволовой части мозга. С другой стороны подходят коллатерали нисходящих моторных трактов, идущих из переднего мозга.
От нейронов ретикулярной формации начинается нисходящий ретикулоспинальный тракт, устанавливающий прямые связи с мотонейронами спинного мозга. Ретикулярная формация имеет связи с ядрами черепно-мозговых нервов, с мозжечком, промежуточным мозгом и через последний - с корой больших полушарий.
Таким образом, богатство связей ретикулярной формации свидетельствует о том, что она является надсегментарной структурой, приспособленной к регуляции деятельности различных отделов центральной нервной системы.
В ретикулярной формации заднего мозга сосредоточены центры, играющие важную роль в регуляции висцеральных функций. Это в первую очередь дыхательный центр, локализованный в ретикулярной формации продолговатого мозга.
Отличительной чертой нейронов дыхательного центра является способность к автоматизму.
В регуляции функций дыхательного центра могут принимать участие и более высокие этажи головного мозга. Так, например, эмоциональные реакции человека связаны с изменением периодичности в работе дыхательного центра, вызванным импульсацией из промежуточного мозга и лимбической коры. Регулирующие воздействия коры головного мозга обеспечивают произвольную регуляцию дыхания, его коррекцию при разнообразных изменениях жизнедеятельности.
Таким образом, регуляция дыхательной функции осуществляется иерархической системой нервных центров, расположенных на разных | этажах нервной системы и связанных единством действия.
Другим жизненно важным центром ретикулярной формации заднего мозга является сосудодвигательный центр. Сосудодвигательный центр занимает обширную область продолговатого мозга.
Стимуляция сосудодвигательного центра вызывает увеличение сосудистого тонуса, повышение кровяного давления и тахикардию.
Рефлекторные влияния на нейроны сосудодвигательного центра осуществляются при возбуждении хемо- и механорецепторов, локализованных в сосудистой стенке.
Для ретикулярной формации стволовой части мозга характерны не только вегетативные регулирующие функции, но и участие в нисходящем контроле деятельности двигательных центров спинного мозга.
Ретикулярная формация как один из двигательных центров стволовой части мозга может выступать не только в роли регулятора возбудимости спинальных мотонейронов, но и принимать участие в процессах, связанных с поддержанием позы и организацией целенаправленных движений.
Наряду с неспецифическими нисходящими влияниями ретикулярной формации ствола были открыты ее восходящие, активирующие влияния на кору головного мозга.
Ретикулярная формация является структурой, отвечающей за состояние бодрствования, структурой, формирующей восходящую активирующую ретикулярную систему, которая поддерживает на определенном уровне возбудимость промежуточного мозга и коры больших полушарий.
Согласно современным представлениям, переход коры к активному состоянию связан с колебаниями количества восходящих сигналов от ретикулярной формации ствола. Количество этих сигналов зависит от поступления в ретикулярную формацию сенсорных импульсов по коллатералям специфических афферентных восходящих путей.
Практически к ретикулярной формации приходит информация от всех органов чувств по коллатералям от спиноретикулярного тракта, проприоспинальных путей, афферентных черепно-мозговых нервов, от таламуса и гипоталамуса, от моторных и сенсорных областей коры.
Средний мозг расположен кпереди от мозжечка и варолиева моста. Морфофункциональная организация среднего мозга. Как видно на поперечном срезе (рис.11.1) дорсальная поверхность среднего мозга занята пластинкой четверохолмия, состоящей из двух пар бугров: верхних и нижних. Верхнее двухолмие играет роль зрительного подкоркового центра и служит местом переключения зрительных путей, идущих к латеральным коленчатым телам промежуточного мозга.

Рис.11.1. Средний мозг (поперечный разрез): тракты 1- кортикомостовой передний, 2 - кортиконуклеарный, 3 - кортикоспинальный латеральный, 4 - кортикоспинальный передний, 5 - кортикомостовой, 6 - сильвиев водопровод, 7 - нижнее двухолмие, 8 - ретикулярная формация, 9 - медиальный лемниск, 10 - латеральный лемниск, 11 - черная субстанция, 12 - красное ядро.
Нижнее двухолмие выполняет функцию подкоркового слухового центра.
В ядрах среднего мозга заложены жизненно важные безусловные рефлексы: сторожевой рефлекс при внезапной подаче светового или звукового раздражителей, рефлекс, вызывающий усиление тонуса мышц сгибателей. В четверохолмии осуществляются замыкание ориентировочных, зрительных и слуховых рефлексов (поворот головы к источнику раздражения, рефлекторная установка на звук внешнего уха), оборонительных рефлексов. Все эти автоматические реакции относятся к категории генетически запрограммированных реакций организма, важных для сохранения вида.
Пигментированная прослойка ножек мозга, состоит из нейронов, богатых пигментом меланином, и носит название черной субстанции. Это образование относится к экстрапирамидной системе регуляции двигательной активности и функционально связано с лежащими в полушариях переднего мозга базальными ганглиями - полосатым телом и бледным шаром.
Одной из структур, ответственных за надсегментарный контроль позного тонуса, является парное красное ядро среднего мозга. Будучи составной частью организующей движения экстрапирамидной системы, красное ядро получает входы от моторной коры, ядер мозжечка, черной субстанции среднего мозга и дает начало руброспинальному тракту, который как общий путь обеспечивает регуляцию тонуса скелетных мышц.
Среднему мозгу принадлежит важная роль в регуляции глазных движений. В рефлекторной регуляции глазных движений принимают участие ядра глазодвигательного (III пара) и блокового (IV пара) черепно-мозговых нервов, расположенные в покрышке. С помощью этого двигательного аппарата глаза могут производить горизонтальные, вертикальные и вращательные движения.
В естественных условиях красное ядро регулирует протекание проприоцептивных рефлексов спинного мозга, подчиняя их рефлексам более высокого порядка.
Средний мозг является не только местом замыкания многих жизненно важных рефлексов, но и выполняет существенную проводниковую функцию. Основание ножек состоит исключительно из нисходящих путей, соединяющих кору больших полушарий с мостом и спинным мозгом. В их числе находятся оба пирамидных тракта, по которым распространяются прямые влияния коры на мотонейроны спинного мозга.
Мозжечок. Структурная организация и связи мозжечка. Мозжечок - крупный вырост варолиева моста, состоящий из двух полушарий и непарного отдела - червя. Со стволовой частью мозга мозжечок соединяется тремя парами ножек. Самые толстые средние ножки как бы охватывают продолговатый мозг и, расширяясь, переходят в варолиев мост. Верхние ножки начинаются в зубчатых ядрах мозжечка (см. ниже) и направляются к четверохолмию среднего мозга. Третья пара ножек (нижняя) спускается вниз, сливаясь с продолговатым мозгом.
Афферентные волокна, приходящие в мозжечок, преимущественно входят в состав средних и нижних ножек, тогда как эфферентные собраны главным образом в верхних ножках мозжечка.
Полушария и червь мозжечка состоят из лежащего на периферии серого вещества - коры и расположенного глубже белого вещества, в котором заложены скопления нервных клеток, образующие ядра мозжечка.
Афферентный вход к нейронному аппарату коры осуществляется через волокна, идущие из продолговатого мозга, от ядер моста, среднего мозга.
В белом веществе мозжечка сконцентрированы три пары ядер. В белом веществе червя близко к срединной плоскости находится ядро шатра. Нейроны этого ядра посылают свои отростки к ретикулярной формации продолговатого мозга и Варолиева моста, где берет свое начало ретикулоспинальный тракт спинного мозга. Латеральнее находится промежуточное ядро, которое разделяется на шаровидное и пробковидное ядра, аксоны которых идут в средний мозг.
Латеральнее всех ядер лежит наиболее крупное зубчатое ядро мозжечка, от которого волокна направляются к таламусу, и далее в моторные зоны коры.
Функции мозжечка. Мозжечок входит в систему регуляции движений, выполняет следующие важные функции: 1) регуляция позы и мышечного тонуса; 2) сенсомоторная координация позных и целенаправленных движений; 3) координация быстрых целенаправленных движений, осуществляемых по команде из коры, больших полушарий.
Эти функции мозжечка наиболее удобно разбирать в соответствии с топической классификацей его отделов, основанной на характере эфферентных связей.
Мозжечок в наибольшей степени связан с управлением тонуса, позы и равновесия тела. В эту зону коры мозжечка, поступает информация, сигнализирующая о позе и состоянии локомоторного аппарата. После обработки этой информации из коры мозжечка через ядро шатра корректирующие команды направляются к ретикулярной формации ствола и оттуда к спинальным центрам
Таким образом, вся система работает по принципу обратной связи и обеспечивает срочную регуляцию антигравитационного тонуса.
Кора мозжечка, связанная со вставочным ядром (или шаровидным и пробковидным), так же имеет афферентные входы от спинальных трактов. Однако, кроме того, через ядра моста кора мозжечка получает информацию от двигательной области коры больших полушарий и сигнализирует о готовящемся целенаправленном движении.
Сопоставление приходящей по этим двум путям информации позволяет мозжечку участвовать в координации целенаправленных движений с рефлексами поддержания позы, в выборе наиболее оптимальной для выполнения движения позы.
Кора полушарий мозжечка получает афферентный вход от различных ассоциативных зон коры больших полушарий. По этим путям в кору полушарий мозжечка поступает информация о замысле движения. В полушариях мозжечка эта информация преобразуется в программу движения, которая поступает в двигательные зоны коры больших полушарий.
Сложная система регуляции движений с наличием обратных связей между мозжечком и корой больших полушарий позволяет полушариям мозжечка участвовать в организации быстрых целенаправленных движений, протекающих без учета информации, идущей от восходящих сенсорных путей спинного мозга. Такие движения могут встретиться в спортивной практике, при игре на музыкальных инструментах и при некоторых других видах деятельности.
Мозжечок участвует не только в регуляции движений, но и включен в систему контроля висцеральных функций: сердечно-сосудистой деятельности, дыхания, моторики и секреторной функции желудочно-кишечного тракта.