

2.2.2. Проприоцептивные ощущения
Изучавшие проблему проприоцепции физиологи не сразу пришли к современным представлениям, согласно которым для формирования проприоцептивпых ощущений нужна информация, поступающая одновременно от разных рецепторов. Она приходит от мышечных веретен и от сухожильных рецепторов Гольджи, от механорецепторов суставных сумок, а также от кожных рецепторов, реагирующих на различную степень растяжения кожи во время движений. Детально исследовавший кожную механорецепцию Макс фон Фрей придавал ей большое значение в проприоцептивном восприятии. Сделав себе кокаиновую анестезию предплечья, он обнаружил, что без зрительного контроля не может правильно поднести вилку ко рту или же прикоснуться п&тьцем к глазу. Таким образом, проприоцептивные ощущения возникают в результате параллельной переработки разных видов информации с последующим синтезом всех поступивших сигналов в соматосенсорной коре.
2.3. Переработка информации в соматосенсорной коре
Как уже было сказано в разделах 2.1 и 2.2, разные виды механорецепторов реагируют на различные характеристики действующих стимулов, тем самым разделяя информацию о них на некоторые элементарные составляющие. Такое разло-
жение целого на части обусловлено особенностями каждого типа рецепторов и демонстрирует самое начало анализа сенсорной информации. От каждого типа рецепторов возбуждение распространяется по собственным проводящим путям параллельно, при этом в каждом переключательном ядре осуществляется независимая переработка сигналов. В результате каждый информационный поток поступает к определенной области соматосенсорной коры, где из разрозненных характеристик стимула должен произойти синтез целостного ощущения.
2.3.1. Топографическая организация соматосенсорной коры
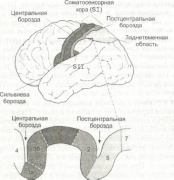
Еще в первой половине XX столетия сначала у животных, а затем у человека было обнаружено топографическое соответствие между различными частями тела и поверхностью мозговой коры. Канадский нейрохирург Уайлдер Пен-филд (Penfield W.) во время проводимых под местной анестезией хирургических операций на человеческом мозге обнаружил, что при электрической стимуляции разных участков постцентральных извилин люди ощущают прикосновение к различным частям противоположной половины тела. Обобщающим результатом этих исследований стало создание карты соматосенсорного представительства в коре (рис. 2.5). Она

64
3 Зак. 1998
65
не соответствует пропорциям человеческого тела, что обусловлено неодинаковой плотностью чувствительной иннервации в разных его частях. Особенно обильно иннервируемые лицо, язык, губы и кисти рук имеют в соматосенсорной коре большую площадь представительства, что обеспечивает этим частям тела способность к наиболее тонкому различению тактильных и проприоцептивных ощущений. Искажение пропорций тела в его представительстве в коре послужили поводом к тому, чтобы определить такое отображение, как сенсорный гомункулус, то есть человечек.
2.3.2. Регионы соматосенсорной коры
Первоначальная схема Пенфилда была уточнена в ходе исследований коры с помощью точечных микроэлектродов, которые позволяли регистрировать активность отдельных нейронов при раздражении ограниченных участков поверхности тела или стимуляции афферентных нейронов, передающих информацию от проприоцепторов. Применение такой техники исследований привело к представлению о существовании функционально различающихся регионов соматосенсорной коры. Эти регионы соответствуют За, 36, 1 и 2 полям Бродмана (рис. 2.6). В поле За поступает информация преимущественно от проприоцепторов мышечных веретен, сухожилий и суставных сумок, в поле 36 — от механорецеп-торов кожи, причем и в том и в другом случае она содержит только элементарные характеристики действующих стимулов. Дальнейшая переработка информации, доставленной от рецепторов кожи, происходит в поле 1, а в поле 2 переработанные сигналы комбинируются с информацией, поступившей от проприоцепторов. Таким образом, если в полях За и 36 формируются только элементарные ощущения действующих раздражителей, то в полях 1 и 2 создаются комплексные представления.
Все четыре поля получают информацию от одной и той же поверхности тела, но в каждом из полей одно ощущение доминирует над остальными. В поле За преобладает вход от интрафузальных рецепторов, в поле 36 — от поверхностных 1рецепторов кожи, в поле 2 — от рецепторов, реагирующих на сильное давление, а в поле 1 — от быстро адаптирую-
Поперечный разрез постцентральной извилины
Рис. 2.6. Регионы соматосенсорной коры в области представительства руки
Опознание предметов на ощупь основано на использовании афферентной информации от тактильных рецепторов (поступает в регион 36) и рецепторов интрафузальных волокон (поступает в регион За). В этих регионах происходит элементарная обработка информации, в которой участвуют простые нейроны коры. Следующий этап переработки информации осуществляется в регионах 1 и 2, где имеются комплексные нейроны, получающие информацию из регионов За и 36 и способные объединять несколько элементарных признаков. Целостное восприятие действующего стимула происходит при участии регионов 5 и 7 заднетеменной области.
щихся рецепторов кожи. В полях За и 36 отсутствуют нейроны, воспринимающие информацию о направлении действия стимула; они содержатся в полях 1 и 2, где определяется трехмерная форма предмета и направление, в котором пере-
66
67
мешается действующий на кожу раздражитель. От некоторых нейронов полей За и 36 аксоны направляются к полям 1 и 2, где конвергируют на одних и тех же клетках, что позволяет последним реагировать на различные комплексные признаки раздражителя, например, на его контур.
Благодаря конвергенции различных афферентных входов к нейронам 1 и 2 регионов их рецептивные поля оказываются большими по сравнению с нейронами За и 36 регионов. Так, например, если рецептивные поля нейронов в регионах За и 36 включают поверхность только одного пальца и один или два сустава, то в регионах 1 и 2 рецептивные поля нейронов благодаря конвергенции от клеток За и 36 регионов включают уже несколько пальцев. Таким образом, обработка информации в соматосенсорной коре происходит последовательно: сначала возбуждается множество нейронов, реагирующих на элементарные признаки стимула, а уже они заставляют активироваться меньшее количество нейронов, предназначенных для интеграции элементарных признаков в комплекс. При ощупывании какого-либо предмета обычно происходит объединение тактильных и проприоцептивных ощущений, то есть сведений о характере поверхности и ее трехмерной форме. Если ощупываемый предмет хорошо знаком человеку (например, монета, расческа, часы), то испытуемый его называет, основываясь на сопоставлении сенсорных ощущений с имеющимся у него опытом. Способность создавать комплексное представление о предмете основана на т.н. стереогностическом чувстве.
Клетки коры, специализирующиеся на переработке однородной информации, объединены в кортикальные колонки, диаметр которых составляет приблизительно 0,2—0,5 мм. Аксоны таламических нейронов, передающих сигналы к коре, в основном заканчиваются в пределах одной колонки, что позволяет ей получать обособленную часть информации и играть роль элементарной функциональной единицы. Принципиальная схема соединений между нейронами разных уровней, обеспечивающая соматопическое представительство в коре того или иного участка тела, определяется генетически, однако индивидуальный сенсорный опыт, в особенности приобретаемый в раннем возрасте, способен вносить в
68
ее изменения. При некоторых обстоятельствах представи-ельство разных частей тела в коре может изменяться и у зрослых животных и людей (справка 2.1).
2.4. Температурная чувствительность
Ощущения тепла или холода, вызванные прикосновени-м к телу человека нагретых или холодных предметов, теп-ого или охлажденного воздуха, возникают вследствие раздражения одной из двух разновидностей температурных рецепторов, представленных в коже свободными нервными окончаниями. Среди них различают тепловые и холодовые рецепторы, которым соответствуют две субмодальности температурного восприятия внешних воздействий. Температурные рецепторы распределены в коже неравномерно и образуют специфические тепловые и холодовые точки площадью около 1 мм2 и менее, с наибольшей плотностью на лице и ладонях, причем Холодовых точек примерно в десять раз больше, чем тепловых.
Тепловые и холодовые точки нетрудно обнаружить с помощью заполненных ледяной водой или водой, подогретой до 45°, конусообразных термодов Бликса (Blix M.), концы которых имеют площадь величиной с булавочную головку. Если легко и быстро прикасаться термодами к разным участ-ам кожи, то в одних случаях возникнет ощущение тепла, а других — холода. Самая высокая плотность температурных рецепторов найдена на лице и ладонях, причем повсюду Холодовых точек обнаруживается примерно в десять раз больше, чем тепловых.
2.4.1. Характеристика Холодовых и тепловых
ТЕРМОРЕЦЕПТОРОВ
Холодовые терморецепторы, гистологическое строение которых окончательно не установлено (предполагают, что они представлены свободными нервными окончаниями), располагаются в эпидермисе и связаны с тонкими миели-низированными волокнами. Тепловые рецепторы, тоже являющиеся, скорее всего, свободными нервными окончаниями, размещены глубже, преимущественно в верхнем и среднем слоях кожи, и соединяются с безмякотными волокнами.
69
О свойствах терморецепторов можно судить по характеру импульсной активности в отходящих от них сенсорных волокнах. У человека приходится использовать для этого вольфрамовые электроды с обнаженным от изоляции кончиком, которые вводятся в нервный ствол через кожу. Импульсная активность терморецепторов возрастает в зависимости от направления и скорости изменений температуры поверхности кожи, составляющей в условиях температурного комфорта около 33°С. При снижении температуры кожи растет активность Холодовых рецепторов с максимумом в диапазоне от 26 до 17°, что субъективно расценивается как действие на кожу холода. При повышении температуры кожи от 34 до 45° увеличивается импульсная активность тепловых рецепторов, что субъективно ощущается как возрастающее по интенсивности действие на кожу тепла. При уменьшении температуры кожи ниже 17° и увеличении выше 45° к температурным ощущениям прибавляются болевые.
При неизменной температуре кожи регистрируется тоническая активность температурных рецепторов, которая постепенно снижается, если действующая температура остается прежней. Снижение импульсной активности объясняется адаптацией рецепторов, субъективно ей соответствует ослабление ощущений тепла или холода. В ответ на резкие ступенчатые изменения действующей температуры (температурный контраст) возникает залповая фазическая активность терморецепторов, которая отражает произошедшие изменения. В этом случае характер температурных ощущений зависит от разницы между исходной температурой кожи и температурой действующего на нее раздражителя (справка 2.2). Наличие адаптации терморецепторов наряду с их способностью реагировать залповой активностью на контрастные изменения действующей температуры обусловливает изменения порогов температурной чувствительности. Поэтому порог тепловой чувствительности снижается при пониженной температуре кожи, а порог холо-довой чувствительности — при повышенной температуре.
2.4.2. Термочувствительные нейроны в ЦНС
Кожные терморецепторы позволяют оценить температуру окружающего воздуха или прикасающихся к коже пред-
70
метов. Но, для того чтобы человек мог сказать «мне жарко» или «мне холодно», возбуждения только таких рецепторов недостаточно. Кроме периферических терморецепторов, расположенных в коже, существуют и многочисленные центральные терморецепторы. Много термочувствительных нейронов имеется в спинном мозгу, но особенно много их в переднем гипоталамусе. Некоторые механорецепторы кожи одновременно являются неспецифическими терморецепторами, их также обнаружили в брюшной полости и в мышцах. Афферентная информация от всех разновидностей терморецепторов поступает в гипоталамус, являющийся главным центром терморегуляции. Однако осознаваемые сенсорные впечатления от контакта с окружающим миром определяются, в первую очередь, активностью периферических кожных терморецепторов.