

Акушерство_Савельева Г.М._2000
.pdfП р е а н т р а л ь н ы й ф о л л и к у л . С началом роста примордиальный фолликул прогрессирует до преантральной стадии, а овоцит увеличивается и окружается мембраной, называемой блестящей оболочкой (zona pellucida). Клетки гранулезного эпителия подвергаются размножению, а слой теки образуется из окружающей стромы. Этот рост характеризуется повышением продукции эстрогенов. Клетки гранулезного слоя преантрального фолликула способны синтезировать стероиды трех классов, при этом эстрогенов син тезируется гораздо больше, чем андрогенов и прогестерона.
А н т р а л ь н ы й , ил и в т о р и ч н ы й , ф о л л и к у л . Характеризуется дальнейшим ростом: увеличивается число клеток гранулезного слоя, проду цирующих фолликулярную жидкость. Фолликулярная жидкость накаплива ется в межклеточном пространстве гранулезного слоя и образует полости.
В этот период фолликулогенеза (8—9-й день менструального цикла) отме чается синтез половых стероидных гормонов, эстрогенов и андрогенов.
Согласно современной теории синтеза половых гормонов, в тека-клет- ках синтезируются андрогены — андростендион и тестостерон. Затем андрогены попадают в клетки гранулезного слоя, и в них ароматизируются в эстрогены.
Д о м и н а н т н ы й ф о л л и к у л . Как правило, один такой фолликул образуется из множества антральных фолликулов (к 8-му дню цикла). Он является самым крупным, содержит наибольшее число клеток гранулезного слоя и рецепторов к ФСГ, ЛГ. Доминантный фолликул имеет богато васкуляризированный тека-слой. Наряду с ростом и развитием доминантного преовуляторного фолликула в яичниках параллельно происходит процесс атрезии остальных (90 %) растущих фолликулов.
Доминантный фолликул в первые дни менструального цикла имеет диаметр 2 мм, который в течение 14 дней к моменту овуляции увеличивается в среднем до 21 мм. За это время происходит ШО-кратное увеличение объема фолликулярной жидкости. В ней резко возрастает содержание эстрадиола и ФСГ, а также определяются факторы роста.
О в у л я ц и я — разрыв преовулярного доминантного (третичного) фол ликула и выход из него яйцеклетки. Ко времени овуляции в овоците про исходит процесс мейоза. Овуляция сопровождается кровотечением из раз рушенных капилляров, окружающих те каклетки. Полагают, что овуляция происходит через 24—36 ч после формирования преовуляторного пика эст радиола. Истончение и разрыв стенки преовуляторного фолликула проис ходят под влиянием фермента коллагеназы. Определенную роль играют также простагландины F2a и Е2, содержащиеся в фолликулярной жидкости;
протеолитические ферменты, образующиеся в гранулезных клетках; окситоцин и релаксин.
После выхода яйцеклетки в полость фолликула быстро врастают обра зующиеся капилляры. Гранулезные клетки подвергаются лютеинизации: в них увеличивается объем цитоплазмы и образуются липидные включения. Л Г, взаимодействуя с белковыми рецепторами гранулезных клеток, стиму лирует процесс их лютеинизации. Этот процесс приводит к образованию Желтого тела.
Ж е л т о е т е л о — транзиторная эндокринная железа, которая функци онирует в течение 14 дней независимо от продолжительности менструаль ного цикла. При отсутствии беременности желтое тело регрессирует.
51
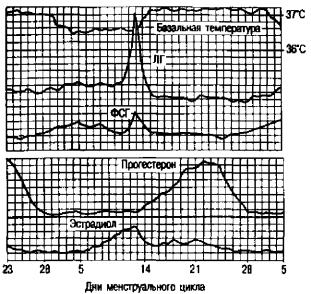
Рис. 2.13. Содержание гормонов в плазме крови в течение менструального цикла.
Таким образом, s яичнике синтезируются основные женские половые стероидные гормоны — эстрадиол и прогестерон, а также андрогены.
ВI фазу менструального цикла, который длится от первого дня менс труации до момента овуляции, организм находится под влиянием эстроге нов, а во II (от овуляции до начала менструации) к эстрогенам присоеди няется прогестерон, выделяющийся кчетками желтого тела. Первая фаза менструального цикла называется также фолликулиновой, или фолликуляр ной, вторая фаза цикла — лютеиновой.
Втечение менструального цикла в периферической крови отмечаются два пика содержания эстрадиола: первый —выраженный преовуляторный цикл, и второй, менее выраженный, —в середине второй фазы менструаль ного цикла. После овуляции во второй фазе цикла основным является прогестерон, максимальное количество которого синтезируется на 4—7-й день после овуляции (рис. 2.13).
Циклическая секреция гормонов в яичнике определяет изменения в слизистой оболочке матки.
Циклические изменения в слизистой оболочке матки (эндометрии). Эндо метрий состоит из следующих слоев.
1.Базальный слой, который не отторгается во время менструации. Из его клеток в течение менструального цикла образуется слой эндо метрия.
2.Поверхностный слой, состоящий из компактных эпителиальных кле ток, которые выстилают полость матки.
3.Промежуточный, или спонгиозный, слой.
Последние два слоя составляют функциональный слой, подвергающий ся основным циклическим изменениям в течение менструального цикла и отторгающийся в период менструации.
52
В I фазе менструального цикла эндометрий представляет собой тонкий слой, состоящий из желез и стромы. Выделяют следующие основные фазы изменения эндометрия в течение цикла:
1)фаза пролиферации;
2)фаза секреции;
3)менструация.
фа з а п р о л и ф е р а ц и и . По мере увеличения секреции эстрадиола растущими фолликулами яичников эндометрий претерпевает пролиферативные изменения. Происходит активное размножение клеток базального слоя. Образуется новый поверхностный рыхлый слой с вытянутыми труб чатыми железами. Этот слой быстро утолщается в 4—5 раз. Трубчатые железы, выстланные цилиндрическим эпителием, удлиняются.
Ф а з а с е к р е ц и и . В лютеиновую фазу яичникового цикла под вли янием прогестерона увеличивается извилистость желез, а просвет их посте пенно расширяется. Клетки стромы, увеличиваясь в объеме, приближаются друг к другу. Секреция желез усиливается. В просвете желез находят обиль ное количество секрета. В зависимости от интенсивности секреции железы либо остаются сильно извитыми, либо приобретают пилообразную форму. Отмечается усиленная васкуляризация стромы. Различают раннюю, сред нюю и позднюю фазы секреции.
М е н с т р у а ц и я . Это отторжение функционального слоя эндометрия. Тонкие механизмы, лежащие в основе возникновения и процесса менструа ции, неизвестны. Установлено, что эндокринной основой начала менструа ции является выраженное снижение уровней прогестерона и эстрадиола вследствие регрессии желтого тела.
Существуют следующие основные локальные механизмы, принимающие
участие в менструации: |
|
1) изменение тонуса спиральных артериол; |
|
2) изменение механизмов гемостаза в матке; |
, |
3)изменения в лизосомной функции клеток эндометрия;
4)регенерация эндометрия.
Установлено, что началу менструации предшествует интенсивное суже ние спиральных артериол, приводящее к ишемии и десквамации эндомет рия.
В течение менструального цикла изменяется содержание лизосом в клетках эндометрия. Лизосомы содержат ферменты, некоторые из которых участвуют в синтезе простагландинов. В ответ на снижение уровня прогес терона усиливается выделение этих ферментов.
Регенерация эндометрия наблюдается с самого начала менструации. К концу 24-го часа менструации отторгается 2/з функционального слоя эндометрия. Базальный слой содержит эпителиальные клетки стромы, яв ляющиеся основой для регенерации эндометрия, которая обычно к 5-му Дню цикла полностью завершается. Параллельно завершается ангиогенез с восстановлением целости разорванных артериол, вен и капилляров.
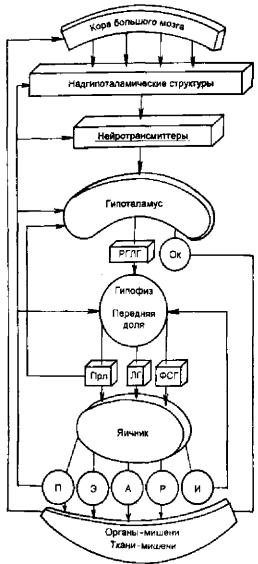
Изменения в яичниках и матке происходят под влиянием двухфазной
Деятельности регулирующих менструальную функцию систем: кора большо го мозга, гипоталамус, гипофиз. Таким образом, выделяются 5 основных звеньев репродуктивной системы женщины: кора большого мозга, гипота ламус, гипофиз, яичник, матка (рис. 2.14). Взаимосвязь всех звеньев репро-
5 3
|
дуктивной системы |
обеспечива |
||||
|
ется наличием в них рецепторов |
|||||
|
как к половым, так и к гонадо- |
|||||
|
тропным |
гормонам. |
|
|
||
|
|
О роли ЦНС в регуляции |
||||
|
функции репродуктивной систе |
|||||
|
мы известно давно. Об этом сви |
|||||
|
детельствовали нарушения овуля |
|||||
|
ции при различных острых и хро |
|||||
|
нических |
стрессах, |
нарушение |
|||
|
менструального цикла при пере |
|||||
|
мене климатогеографических зон, |
|||||
|
ритма работы; |
хорошо известно |
||||
|
прекращение менструаций в ус |
|||||
|
ловиях военного времени.У пси |
|||||
|
хически неуравновешенных жен |
|||||
|
щин, страстно желающих иметь |
|||||
|
ребенка, менструации также могут |
|||||
|
прекратиться. |
|
|
|
||
|
|
В к о р е |
б о л ь ш о г о |
|||
|
м о з г а |
и в экстрагипоталами- |
||||
|
ческих церебральных структурах |
|||||
|
(лимбической |
системе, гиппо- |
||||
|
кампе, миндалине и др.) выявле |
|||||
|
ны |
специфические |
рецепторы |
|||
|
для |
эстрогенов, прогестерона |
и |
|||
|
андрогенов. В |
этих |
структурах |
|||
|
происходят синтез, выделение и |
|||||
|
метаболизм нейропептидов, ней- |
|||||
|
ротрансмиттеров и их рецепто |
|||||
|
ров, которые в свою очередь из |
|||||
|
бирательно влияют на синтез и |
|||||
|
выделение рилизинг-гормона ги |
|||||
|
поталамуса. |
|
|
|
||
|
|
Во взаимосвязи с половыми |
||||
|
стероидами функционируют ней- |
|||||
|
ротрансмиттеры: норадреналин, |
|||||
|
дофамин, |
гамма-аминомасляная |
||||
Рис. 2.14. Функция репродуктивной системы |
кислота, ацетилхолин, серотонин |
|||||
(схема). |
и мелатонин. Норадреналин сти |
|||||
РГЛГ — рилизинг-гормоны; ОК. — оксито- |
мулирует |
выброс гонадотропин- |
||||
иин; Прл — пролактин, ФСГ — фолликуло- |
рилизинг-гормона |
(ГТРГ) |
из |
|||
стимулирующий гормон; П — прогестерон; |
нейронов переднего |
гипоталаму |
||||
Э — эстрогены; А — андрогены; Р — релак |
са. Дофамин и серотонин умень |
|||||
син; И — ингибин; ЛГ — лютеинизируюший |
шают частоту и снижают ампли |
|
гормон. |
||
туду выработки ГТРГ в различ |
||
|
||
|
ные фазы менструального цикла. |
Нейропептиды (эндогенные опиоидные пептиды, нейропептид Y, кор- тикотропин-рилизинг-фактор и галанин) также влияют на функцию репро дуктивной системы, а следовательно, на функцию гипоталамуса. Эндоген-
54
ные опиоидные пептиды трех видов (эндорфины, энкефалины и динорфины) способны связываться с опиатными рецепторами мозга. Эндогенные опиоидные пептиды (ЭОП) модулируют влияние половых гормонов на содержание ГТРГ по механизму обратной связи, блокируют секрецию ги пофизом гонадотропных гормонов, особенно ЛГ, посредством блокады се креции ГТРГ в гипоталамусе.
Взаимодействие нейротрансмиттеров и нейропептидов обеспечивает в организме женщины репродуктивного возраста регулярные овуляторные циклы, влияя на синтез и выделение ГТРГ гипоталамусом.
В г и п о т а л а м у с е имеются пептидергические нейронные клетки, которые секретируют стимулирующие (либерины) и блокирующие (статины) нейрогормоны — нейросекреция. Эти клетки обладают свойствами как ней ронов, так и эндокринных клеток, и отвечают как на сигналы (гормоны), поступающие из кровотока, так и на нейротрансмиттеры и нейропептиды мозга. Нейрогормоны синтезируются в рибосомах цитоплазмы нейрона, а затем транспортируются по аксонам к терминалям.
Гонадотропин-рилизинг-гормон (либерин) — нейрогормон, регулирую щий гонадотропную функцию гипофиза, где синтезируются ФСГ и ЛГ. Рилизинг-гормон ЛГ (люлиберин) выделен, синтезирован и подробно опи сан. Выделить и синтезировать рилизинг-фолликулостимулирующий гор мон, или фоллиберин, до настоящего времени не удалось.
Секреция гонадолиберина имеет пульсирующий характер: пики усилен ной секреции гормона продолжительностью несколько минут сменяются 1 — 3-часовыми интервалами относительно низкой секреторной активности. Час тоту и амплитуду секреции гонадолиберина регулирует уровень эстрогенов.
Нейрогормон, который контролирует секрецию пролактина аденогипофизом, называется пролактинингибирующим гормоном (фактором), или дофамином.
Важным звеном в системе репродукции является передняя доля ги п о ф и з а — аденогипофиз, в котором секретируются гонадотропные гор моны, фолликулостимулирующий гормон (ФСГ, фоллитропин) лютеинизирующий гормон (ЛГ, лютропин) и пролактин (Прл), регулирующие функ
цию яичников и молочных желез. Все три гормона являются белковыми веществами (полипептидами). Железой-мишенью гонадотропных гормонов является яичник.
В передней доле гипофиза синтезируются также тиреотропный (ТТГ) и адренокортикотропный (АКТГ) гормоны, а также гормон роста.
ФСГ стимулирует рост и созревание фолликулов яичника, способствует образованию рецепторов ФСГ и ЛГ на поверхности гранулезных клеток
яичника, увеличивает содержание ароматаз в зреющем фолликуле и, стиму лируя процессы ароматизации, способствует превращению андрогенов в эстрогены, стимулирует продукцию ингибина, активина и инсулиноподоб ного фактора роста-1, которые играют ингибирующую и стимулирующую
роль в росте фолликулов.
ЛГ стимулирует:
•образование андрогенов в тека-клетках;
•овуляцию совместно с ФСГ;
•ре моделирование гранулезных клеток в процессе лютеинизации;
•синтез прогестерона в желтом теле.
55
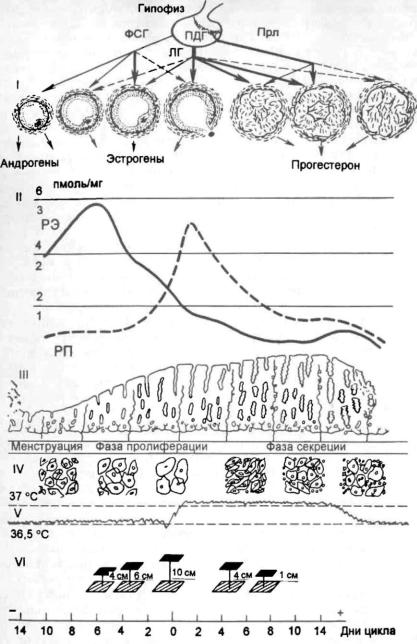
Рис. 2.15. Циклические изменения в органах репродуктивной системы в течение менструального цикла.
I — гонадотропная регуляция функции яичников: ПДГ — передняя доля гипофиза, остальные обозначения те же, что на рис. 2.14; II — содержание в эндометрии рецепторов к эстрадиолу — РЭ (1,2,3; сплошная линия) и прогестерону — РП (2,4,6; пунктирная линия); III — циклические изменения эндометрия; ГУ — цитология эпи телия влагалища; V — базальная температура; VI — натяжение цервикальной слизи.
56
Пролактин стимулирует рост молочных желез и лактацию, контролирует секрецию прогестерона желтым телом путем активации образования в них рецепторов к ЛГ.
Синтез пролактина аденогипофизом находится под тоническим блоки рующим контролем дофамина, или пролактинингибирующего фактора. Ингибиция синтеза пролактина прекращается во время беременности, лакта ции. Основным стимулятором синтеза пролактина является тиролиберин, синтезируемый в гипоталамусе.
Циклические изменения в гипоталамо-гипофизарной системе и в яич никах взаимосвязаны и моделируются по типу обратной связи.
Выделяют следующие типы обратной связи:
1)"длинная петля" обратной связи — между гормонами яичника и ядрами гипоталамуса; между гормонами яичника и гипофизом;
2)"короткая петля" — между передней долей гипофиза и гипоталаму сом;
3)"ультракороткая петля" — между ГТРГ и нервными клетками гипо таламуса.
Взаимосвязь всех указанных структур определяется наличием в них рецепторов к половым гормонам.
Уженщины репродуктивного возраста имеется как отрицательная, так
иположительная обратная связь между яичниками и гипоталамо-гипофи зарной системой. Примером отрицательной обратной связи является усиле ние выделения Л Г передней долей гипофиза в ответ на низкий уровень эстрадиола в раннюю фолликулярную фазу цикла. Примером положитель ной обратной связи является выброс ЛГ в ответ на овуляторный максимум содержания эстрадиола в крови.
Осостоянии репродуктивной системы можно судить по оценке тестов функциональной диагностики: базальная температура, симптом зрачка и кариопикнотеческий индекс (рис. 2.15).
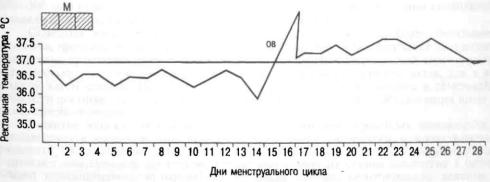
Б а з а л ь н а я т е м п е р а т у р а |
измеряется в прямой кишке утром, до |
|
подъема с постели. При овуляторном менструальном цикле |
базальная тем |
|
пература повышается в лютеиновую |
фазу цикла на 0,4—0,6 |
°С и держится |
в течение всей второй фазы (рис. 2.16). В день менструации или за день до
Рис. 2.16. Ректальная температура при двухфазном цикле. М — менструация; О В — овуляция.
57
нее баэаотьиая чнййяературй'"-снижается. При берешннвйтИ повышение базальной температуры объясняется возбуждением терморегулирующего цент ра гипоталамуса под влиянием прогестерона.
С и м п т о м з р а ч к а отражает изменения в слизи шейки матки. Под влиянием эстрогенов в шейке матки накапливается прозрачная стекловид ная слизь, что обусловливает расширение наружного отверстия шейки матки. Максимальное количество слизи наблюдается в предовуляторные дни цикла, наружное отверстие становится темным, напоминает зрачок. Во вторую фазу цикла под влиянием прогестерона количество слизи уменьша ется или же она полностью исчезает. Слизь имеет глыбчатое строение. Различают 3 степени симптома зрачка: +, + + , + + + .
К а р и о п и к н о т и ч е с к и й и н д е к с . Под влиянием гормонов яич ников также происходят циклические изменения в слизистой оболочке влагалища, особенно в его верхней трети. В мазке из влагалища могут встречаться следующие виды клеток плоского многослойного эпителия: а) ороговевающие, б) промежуточные, в) базальные, или атрофические. Клетки первого типа начинают преобладать по мере нарастания секреции яичниками эстрогенов. На основании определения количественных соотно шений клеточных элементов можно судить о степени насыщенности орга низма эстрогенными гормонами или об их недостаточности. Максимальное число ороговевающих клеток выявляется в предо вуляторные дни — 80—88 %, в раннюю фазу пролиферации — 20—40 %, в позднюю фазу секреции — 20-25 %.
Глава 3 ФИЗИОЛОГИЯ БЕРЕМЕННОСТИ
3.1. ОПЛОДОТВОРЕНИЕ. РАННИЙ ЭМБРИОГЕНЕЗ
3.1.1.Оплодотворение
•Оплодотворением называется процесс слияния зрелых мужской (спер матозоид) и женской (яйцеклетка) половых клеток, в результате чего возникает зигота, несущая генетическую информацию как отца, так
иматери.
Процесс созревания мужских и женских половых клеток очень сложен. Спер матогенез совершается в извитых семенных канальцах мужских гонад. Он заверша ется в период половой зрелости образованием зрелых сперматозоидов, обладающих способностью к оплодотворению. Полному созреванию предшествует процесс редук ционного деления, в результате которого в ядре сперматозоида содержится гаплоид ный набор хромосом.
Сперматозоиды бывают двух видов: носители половых Х- и Y-хромосом. При слиянии с яйцеклеткой сперматозоида, являющегося носителем половой Х-хромо- сомы, из образующейся зиготы развивается эмбрион женского пола, при слиянии сперматозоида, имеющего половую Y-хромосому, возникает зародыш мужского пола (яйцеклетка всегда является носительницей половой Х-хромосомы).
Зрелый сперматозоид имеет длину до 50—60 мкм и состоит из головки, шейки и хвостовой части. Головка сперматозоида, имеющая овальную форму, содержит ядро, окруженное тонким слоем протоплазмы. Шейка имеет протоплазму, содержа щую видоизмененную центросому, которая способствует процессу дробления опло дотворенной яйцеклетки. Хвостик состоит из протоплазмы и выполняет двигатель ные функции, В результате колебательных движений хвостовой части сперматозоиды способны совершать самостоятельные движения в половом тракте женщины со скоростью 2—3 мм/мин. Способность к движениям сперматозоиды получают после их контакта с секретом семенных пузырьков и предстательной железы. Такая смесь носит название семенной жидкости, или спермы. При половом сношении во влага лище женщины изливается в среднем около 3—5 мл спермы, в которой содержится 300—500 млн сперматозоидов. Использование электронной микроскопии позволило установить весьма сложную структуру сперматозоида (рис. 3.1).
Семенная жидкость имеет сложный состав и содержит фруктозу, белковые вещества, протеазы, кислую фосфатазу, лимонную кислоту и биологически активные вещества — простагландины. Семенная жидкость обладает выраженной антигенной активностью, при этом антигены обнаружены как в самих сперматозоидах, так и в жидкой части спермы. Полагают, что число антигенных компонентов в семенной жидкости достигает 12. В процессе оплодотворения происходит нейтрализация анти генов сперматозоидов.
Развитие яйцектеток связано с ростом и развитием первичных фолликулов, находящихся в корковом слое яичников. Созревание яйцеклетки условно можно разделить на созревание ядра и созревание цитоплазмы. Под созреванием ядра понимают совокупность процессов, переводящих ядро из стадии динлотена I (или стадии герминативного пузырька) до метафазы второго мейотического деления. Созревание ядра не включает в себя завершение мейоза, так как только проникно вение сперматозоида внутрь яйцеклетки вызывает завершение второго редукцион ного деления.
59

Рис .3.1. Строение сперматозоида человека (электронно-микроскопическая схема), ] — акросома; 2 — ядро; 3 — шейка; 4 — митохондрии; 5 — осевые нити.
Рис. 3.2. Зрелая яйцеклетка, окру женная клетками лучистого венца.
От периода наступления полового со зревания до менопаузы у женщины в каждом менструальном цикле обычно созревает одна яйцеклетка. Овоцит первого порядка пре вращается в овоцит второго порядка при от щеплении первого полярного тельца. В мо мент овуляции овоцит второго порядка ока зывается блокированным на стадии метафазы второго мейотического деления. Со зревание яйцеклеток стало известно в дета лях благодаря возможности культивирова ния яйцеклеток in vitro, что в настоящее время широко используют при экстракорпо ральном оплодотворении.
Зрелая яйцеклетка состоит из ядра, ци топлазмы, окружена блестящей оболочкой и клетками лучистого венца, представляющего собой остатки гранулезных клеток фоллику ла (рис. 3.2). Женская половая клетка, как и мужская, обладает антигенными свойствами. Особенно богата различными антигенами ее блестящая оболочка.
Прежде чем анализировать механизмы оплодотворения, т.е. процесс слияния яйцеклетки со сперматозоидом, следует осветить вопросы транс порта гамет. В первую очередь это касается продвижения сперматозоидов по каналам репродуктивной системы женщины. При нормальной эякуляции во влагалище попадает в среднем около 100 млн сперматозоидов, некоторые из которых имеют те или иные морфологические либо функциональные отклонения. Часть сперматозоидов, в том числе и неполноценных, остается во влагалище и подвергается фагоцитозу, Вместе со сперматозоидами во влагалище попадают и другие составные части спермы, при этом особая роль принадлежит проста гланд инам. Под их влиянием происходит актива ция сократительной активности матки и маточных труб, что очень важно для нормального транспорта гамет.
Из влагалища, имеющего у здоровой женщины кислую среду (кислая
60