

Вопрос 19. Система сигнализации: эндокринная, синоптическая. Роль медиаторов и гормонов.
Шпора 10.
Роль медиаторов: обеспечение механической прочности, обмен вещест, транспорт веществ
Вопрос 20. Понятие о вторичных посредниках. Инозитолфосфатная система
Функционирование инозитолфосфатной системы трансмембранной передачи сигнала обеспечивают: R (рецептор), фосфолипаза С, Gрlс - белок, активирующий фосфолипазу С, белки и ферменты мембран и цитозоля.
Последовательность событий, приводящих к активации фосфолипазы С:
связывание сигнальной молекулы, например гормона с рецептором (R) вызывает изменение конформации и увеличение сродства к Gplc-белку.
образование комплекса [Г] [R] [Gрlс ГДФ] приводит к снижению сродства α-протомера G рlс белка к ГДФ и увеличению сродства к ГТФ. ГДФ заменяется на ГТФ.
это вызывает диссоциацию комплекса; отделившаяся α-субъединица, связанная с молекулой ГТФ, приобретает сродство к фосфолипазе С.
α-ГТФ взаимодействует с фосфолипазой С и активирует её. Под действием фосфолипазы С происходит гидролиз липида мембраны фосфатидилинозитол-4,5 - биофосфата (ФИФ2).
в ходе гидролиза образуется и выходит в цитозоль гидрофильное вещество инозитол-1,4,5-трифосфат (ИФ3). Другой продукт реакции диацилглицерол (ДАГ) остаётся в мембране и участвует в активации фермента протеинкиназы С (ПКС).
инозитол-1,4,5-трифосфат (ИФ3) связывается специфическими центрами Са2 - канала мембраны ЭР, это приводит к изменению конформации белка и открытию канала - Са²+ поступает в цитозоль. В отсутствие в цитозоле ИФ3 канал закрыт.
Активация протеинкиназы С.
• Повышение концентрации Са²+ в цитозоле клетки увеличивает скорость
взаимодействия Са²+ с неактивным цитозольным ферментом протеинкиназой С(ПКС) и белком кальмодулином, таким образом сигнал, принятый рецептором клетки, раздваивается.
• Связывание протеинкиназы С с ионами кальция позволяет ферменту вступать в кальций-опосредованное взаимодействие с молекулами "кислого" фосфолипида мембраны, фосфатидилсерина (ФС). Диацилглицерол, занимая специфические центры в протеинкиназе С, ещё более увеличивает её сродство к ионам кальция.
• На внутренней стороне мембраны образуется ферментативный комплекс - [ПКС] [Са²+] [ДАГ] [ФС] - активная протеинкиназа С, фосфорилирующая специфические ферменты по серину и треонину.
Вопрос 21. Понятие о вторичных посредниках. Аденилатцеклазная система
Наиболее изученным является аденилатциклазный путь передачи гормонального сигнала. В нем задействовано мимимум пять хорошо изученных белков: 1)рецепторгормона; 2)ферментаденилатциклаза, выполняющая функцию синтеза циклическогоАМФ(цАМФ); 3) G-белок, осуществляющий связь междуаденилатциклазойирецептором; 4) цАМФ-зависимаяпротеинкиназа, катализирующаяфосфорилированиевнутриклеточныхферментовили белков-мишеней, соответственно изменяя ихактивность; 5)фосфодиэстераза, которая вызывает распадцАМФи тем самым прекращает (обрывает) действие сигнала (рис. 8.5).
Получены в чистом виде α- и β-адренергические рецепторыизплазматических мембранклетокпечени, мышц ижировой ткани. Показано, что связываниегормонас β-адренергическимрецепторомприводит к структурным изменениям внутриклеточногодоменарецептора, что в свою очередь обеспечивает взаимодействиерецепторасо вторымбелкомсигнального пути – ГТФ-связывающим.
ГТФ-связывающий белок– G-белок – представляет собой смесь 2 типовбелков: активного Gs(от англ. stimulatory G) и ингибиторного Gi с мол. массой 80000–90000. В составе каждого из них имеется три разные субъединицы (α-, β- и γ-), т.е. это гетеротримеры. Показано, что β-субъеди-ницы Gsи Giидентичны (мол. масса 35000); в то же время α-субъединицы, являющиеся продуктами разныхгенов(мол. масса 45000 и 41000), оказались ответственными за проявление G-белком активаторной и ингибиторнойактивностисоответственно. Гормонрецепторный комплекс сообщает G-белку способность не только легко обменивать эндогенный связанный ГДФ на ГТФ, но и переводить Gs-белок в активированное состояние, при этом активный G-белок диссоциирует в присутствииионовMg2+на β-, γ-субъединицы и комплекс α-субъединицы Gsв ГТФ-форме; этотактивный комплексзатем перемещается кмолекулеаденилатциклазыи активирует ее. Сам комплекс затем подвергается самоинактивации за счет энергии распада ГТФ и реассоциации β- и γ-субъединиц с образованием первоначальной ГДФ-формы Gs.
Вопрос №22 межклеточные контакты. Адгезивне соединения.
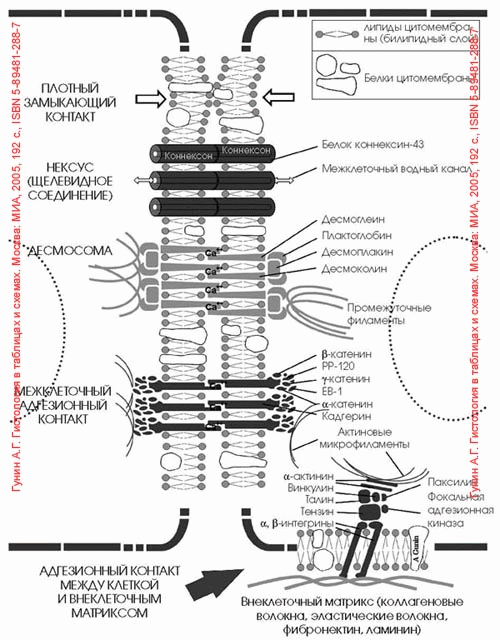
Межклеточные точечные сцепляющие соединения обнаружены у многих неэпителиальных тканей, но более отчетливо описана структура специальных (адгезивных) лентв однослойных эпителиях. Это структура опоясывает весь периметр эпителиальной клетки, подобно тому как это происходит в случае плотного соединения. Чаще всего такой поясок или лента лежит ниже плотного соединения. В этом месте плазматические мембраны не сближены, а даже несколько раздвинуты на расстояние 25-30 нм, и между ними видна зона повышенной плотности. Это ничто иное как места взаимодействия трансмембранных гликопротеидов, которые специфически сцепляются друг с другом и обеспечивают механическое соединение мембран двух соседних клеток. Эти линкерные белки относятся к Е-кадгеринам - белкам, обеспечивающим специфическое узнавание клетками однородных мембран. Разрушение этого слоя гликопротеидов приводит к обособлению отдельных клеток и разрушению эпителиального пласта. С цитоплазматической стороны около мембраны видно скопление какого-то плотного вещества, к которому примыкает слой тонких (6-7 нм) филаментов, лежащих вдоль плазматической мембраны в виде пучка, идущего по всему периметру клетки. Тонкие филаменты относятся к актиновым фибриллам, они связываются с плазматической мембраной посредством белка катенина, образующего плотный около мембранный слой.
Функциональное значение такого ленточного соединения заключается на только в механическом сцеплении клеток друг с другом: при сокращении актиновых филаментов в ленте может изменяться форма клетки. Считается, что кооперативное сокращение актиновых фибрилл во всех клетках эпителиального пласта может вызвать изменение его геометрии, например, сворачивание в трубку, подобно тому, что происходит при образовании нервной трубки у эмбрионов позвоночных.
Шпора 10.
Вопрос №23.межклеточные соединения. Проводящие (химические)контакты
Нексусы (щелевидные контакты):
Контакт образуется на небольшом по площади участке.
В месте контакта в цитомембрану встроены трансмембранные белки коннексины, которые соединяются между собой и образуют водный канал в толще мембраны — конексон.
Коннексоны контактирующих клеток соединяются (или сопоставляются), в результате чего между соседними клетками образуется канал, с помощью которого из одной клетки в другую (в обоих направлениях) свободно проходит вода, малые молекулы и ионы, а также электрический ток.
Синапсы — см. раздел по нервной ткани. Обеспечивают передачу потенциала действия (нервного импульса) с нервной клетки на другую нервную или иную клетку.
Шпора 10