

2 курс / Нормальная физиология / Общий_курс_физиологии_человека_и_животных_Том_2_Ноздрачев_А_Д_,
.pdfпроникает из межклеточной жидкости в просвет прямого сосуда и тонкого отдела петли Генле.
Поднимаясь вместе с током жидкости по направлению к коре почки по прямому сосуду, мочевина непрерывно участвует в противоточном обмене, диффундирует в нисходящий отдел прямого сосуда и нисходящую часть петли Генле. Вследствие этого происходит постоянное поступление мочевины во внутреннее мозговое вещество, а также ионов Сl- и Na+, реабсорбируемых клетками толстого восходящего отдела петли Генле и собирательных трубок. Эти вещества удерживаются в мозговом веществе благодаря деятельности противоточной системы прямых сосудов и петель Генле, что, в конечном счете, обеспечивает повышение осмотической концентрации во внутреннем мозговом веществе почки. Вслед за увеличением осмолярности межуточной ткани, окружающей собирательные трубки, возрастает и реабсорбция воды из них, повышается эффективность саморегулирующей функции почки. Увеличение проницаемости канальцевой стенки для мочевины в присутствии АДГ позволяет понять, почему при снижении мочеотделения уменьшается очищение от мочевины.
Прямые кровеносные сосуды мозгового вещества почки, подобно канальцам петли Генле, также образуют противоточную систему, играющую очень важную роль в осмотическом концентрировании. Благодаря параллельному расположению прямых сосудов и петель Генле обеспечивается эффективное кровоснабжение мозгового вещества почки, но не происходит вымывания осмотически активных веществ, так как в крови прямых сосудов наблюдаются такие же изменения осмотической концентрации, как и в тонком нисходящем отделе петли Генле. При движении крови по направлению к вершине почечного сосочка в ней постепенно возрастает осмотическая концентрация, а во время ее обратного движения к коре почки, диффундирующие через сосудистую стенку соли и другие растворенные вещества переходят в межклеточную ткань.
Тем самым сохраняется градиент концентрации осмотически активных веществ внутри почки, и прямые сосуды функционируют как противоточная система. Скорость движения крови по прямым сосудам влияет на количество удаляемых из мозгового вещества ионов натрия, хлора и мочевины, участвующих в создании осмотического градиента и оттоке реабсорбируемой воды.
Для количественной оценки концентрационной способности почки в условиях экономии воды используют два параметра: индекс осмотического концентрирования,
показывающий, во сколько раз моча более осмотически концентрирована, чем кровь, и
объем реабсорбированной осмотически свободной воды. Физиологическое значение этого показателя, имеющего очень важное значение; требует специального объяснения.
Образование мочи всегда происходит из жидкости, имеющей такую же общую концентрацию осмотически активных веществ, как и плазма крови. Так как выделение почкой осмотически активных веществ равно произведению диуреза (V, мл/мин) на концентрацию осмотически активных веществ в моче (UОsm, мосм/л), то очищение плазмы крови от осмотически активных веществ (CОsm) равно CОsm = UОsmV/PОsm, где PОsm — концентрация осмотически активных веществ в плазме крови, мосм/л. Величина CОsm характеризует тот условный объем плазмы крови, который очистился от осмотически активных веществ в течение 1 мин. Иными словами, если выделяется моча, имеющая такую же осмотическую концентрацию, как и плазма крови, то CОsm = V и почка только уменьшает объем жидкости в организме, не участвуя в осморегуляции.
Однако при обезвоживании почка выделяет мочу более осмотически концентрированную, чем кровь. Воду, выделяемую с мочой, условно можно разделить на две фракции. Одна из них содержит растворенные вещества в той же концентрации, что и плазма крови, т. е. равна CОsm, другая представляет собой чистую, свободную от веществ воду. Ее называют осмотически свободной водой (СH2O ). Следовательно, сказанное можно
представить в виде такой формулы: V = CОsm + СH2O .
Образование гипотонической мочи. При потреблении больших количеств воды
почки выделяют гипотоническую мочу и СH2O представляет собой положительную
величину, при обезвоживании организма, напротив, отрицательную. Это означает, что осмотически свободная вода не экскретируется, а всасывается в канальцах в кровь. Выше был подробно описан механизм осмотического концентрирования мочи. Величина реабсорбции (мл/мин) осмотически свободной воды (Тc H2O ) численно равна СH2O , но с
обратным знаком: Тc H2O = CОsm — V. Существенно, что эта величина является
константной для данного вида. Так, у человека при дегидратации максимальное значение реабсорбции осмотически свободной воды не превышает 5 мл/мин при нормальном значении клубочковой фильтрации.
При поступлении избытка воды в организм почки начинают выделять большие объемы гипотонической мочи. Переход от экономии к экскреции воды сопровождается снижением клубочковой фильтрации в юкстамедуллярных нефронах и ее увеличением в клубочках других популяций нефронов.
При водном диурезе относительная проксимальная реабсорбция ионов и воды не изменяется, а в дистальный сегмент нефрона поступает такое же количество жидкости, как и при дегидратации. Осмолярность мозгового вещества почки при водном диурезе становится ниже, чем при антидиурезе, а осмотическая концентрация жидкости, поступающей в дистальный сегмент нефрона, такая же — приблизительно 200 мосм/л. При водном диурезе стенка конечных отделов почечных канальцев остается водонепроницаемой, а из протекающей мочи клетки продолжают реабсорбировать соли натрия; в результате выделяется гипотоническая моча, концентрация осмотически активных веществ в которой ниже 50 мосм/л. Проницаемость канальцев для мочевины низка, и она экскретируется с мочой, не накапливаясь в мозговом веществе почки.
У человека на максимуме водного диуреза мочеотделение может достигать 18 мл/мин. Очищение от осмотически свободной воды в этом случае определяется по формуле СH2O
= V — Cosm, оно составляет у человека 13—15 мл/мин.
Таким образом, различие деятельности петли Генле, конечных частей дистального сегмента и собирательных трубок обусловливает способность почек при водной нагрузке выделять большие объемы резведенной, гипотонической, мочи, а при дефиците воды в организме экскретировать малые количества мочи, осмотически более концентрированной, чем кровь. Исключительно развита способность почки осмотически концентрировать мочу у некоторых пустынных грызунов, что позволяет им вообще не пить воду. У большой песчанки, например, концентрация осмотически активных веществ в моче может достигать 4000—4500 мосм/кг Н2О, у ряда видов пустынных мышей она возрастает до 9000 мосм/кг Н2О и выше.
Собирательные трубки обеспечивают реабсорбцию Na+, Сl- и других ионов против высокого градиента. Их основная функциональная особенность состоит в том, что реабсорбция веществ происходит в небольших количествах, но против наиболее значительного градиента, что и обусловливает резкие различия концентрации ряда неорганических веществ в моче по сравнению с кровью.
12.6.7. Роль почек в осморегуляции и волюморегуляции
Осморегуляция. Почки являются основным эффекторным органом системы осморегуляции. Они обеспечивают выделение избытка воды в виде гипотонической мочи при поступлении воды в организм или экономят воду и экскретируют мочу, гипертоническую по отношению к плазме крови, при обезвоживании. При избыточном содержании воды в организме концентрация растворенных осмотически активных веществ в крови снижается и ее осмотическое давление падает. Это уменьшает активность центральных осморецепторов, расположенных в области супраоптического ядра гипоталамуса, а также периферических осморецепторов, имеющихся в печени, почке,

селезенке и ряде других органов. Уменьшение активности осморецепторов снижает секрецию антидиуретического гормона (АДГ) нейрогипофизом и приводит к возрастанию выделения воды почкой (рис. 12.14).
Рис. 12.14. Участие почки в регуляции водно-солевого обмена: 1 — ЦНС, 2 — гипоталамус, 3 —
нейрогипофиз, 4 — почка, 5 — поджелудочная железа, 6 — надпочечник; пунктирными стрелками обозначено влияние гормонов, регулирующих обмен ионов
При обезвоживании организма при введении в сосудистое русло гипертонического раствора хлористого натрия увеличивается концентрация осмотически активных веществ в плазме крови, возбуждаются осморецепторы, стимулируются нейроны супраоптического ядра, усиливается секреция АДГ .возрастает реабсорбция воды в канальцах, уменьшается мочеотделение и выделяется осмотически концентрированная. моча. Установлено, что у человека в норме при осмоляльности крови около 280 мосм/кг Н2О полностью прекращается секреция АДГ и почка выделяет наибольшие количества воды. При обезвоживании, когда осмоляльность крови возрастает до 295 мосм/кг Н2О, секреция АДГ достигает максимума и обнаруживается пик осмотического концентрирования мочи в почке. В эксперименте показано, что секреция АДГ возрастает при раздражении не только осморецепторов, но и специфических натриорецепторов. После введения в область III желудочка мозга гипертонического раствора NaCl наступает антидиурез, а после введения в ту же область гипертонических растворов сахара мочеотделение не уменьшается.
Осморецепторы реагируют на сдвиги концентрации осмотически активных веществ в плазме крови. При увеличении осмоляльности плазмы крови на 1 % (около 3 мосм/л) концентрация аргинин-вазопрессина, являющегося антидиуретическим гормоном у человека и многих других млекопитающих, возрастает на 1 пг/мл плазмы. Повышение концентрации осмотически активных веществ в плазме на 1 мосм/кг воды вызывает благодаря выделению АДГ увеличение осмотической концентрации мочи почти в 100 раз, а переход от состояния водного диуреза до максимального осмотического концентрирования мочи требует всего 10-кратного возрастания АДГ в крови — с 0,5 до 5 пг/мл.
Волюморецепция. Помимо информации от осморецепторов и натриорецепторов уровень секреции АДГ зависит и от активности волюморецепторов, реагирующих на изменение объема внутрисосудистой и внеклеточной жидкости. Ведущее значение в регуляции секреции АДГ имеют те волюморецепторы, которые реагируют на изменение напряжения сосудистой стенки. В первую очередь это рецепторы левого предсердия, импульсы от которых передаются в ЦНС по афферентным волокнам блуждающего нерва. При увеличении кровенаполнения левого предсердия активируются волюморецепторы и угнетается секреция АДГ, тем самым усиливается мочеотделение. В предсердии вырабатывается гормон, увеличивающий выделение ионов Na почкой и получивший
название атриальный натрийуретический пептид.
Рассмотрим особенности реакции почки при возбуждении волюморецепторов и осморецепторов. Активация только волюморецепторов возникает в ответ на увеличение объема внутрисосудистой жидкости при неизмененной концентрации в ней осмотически активных веществ. Это приводит к повышению экскреции солей натрия и воды. Одновременная активации волюморецепторов и осморецепторов может наступить при потреблении больших количеств воды, в этом случае в основном усиливается выведение воды. Ведущую роль в изолированном осморегулирующем рефлексе играет уменьшение секреции АДГ, в результате снижается реабсорбция воды и усиливается ее экскреция.
Увеличение объема внеклеточной жидкости, возрастание объема крови служат стимулом для волюморегулирующего рефлекса. В эксперименте адекватным разражителем системы регуляции объема внеклеточной жидкости может служить введение в
кровеносный сосуд больших количеств изотоничного плазме крови физиологического раствора. Возрастание притока крови к сердцу, растяжение стенки предсердия стимулируют волюморецепторы и секрецию из клеток предсердия атриального натрийуретического пептида. Под влиянием этого гормона усиливается выделение ионов натрия и воды почкой.
Стимуляция волюморецепторов рефлекторно изменяет характер импульсации иннервирующих почку симпатических нервов и снижает реабсорбцию натрия и воды в канальцах. Возрастание объема крови способствует растяжению афферентной артериолы клубочка. Это сопровождается изменением активности юкстагломерулярного аппарата, уменьшением секреции фермента ренина, благодаря чему снижается образование в крови из ангиотензиногена ангиотензина I. После отщепления от него двух аминокислот образуется ангиотензин II, который вызывает ряд эффектов, в том числе стимулирует секрецию альдостерона. Когда снижается количество ангиотензина, падает и поступление в кровь альдостерона, что приводит к уменьшению реабсорбции натрия в канальцах. Активация волюморегу-лирующего рефлекса снижает секрецию вазопрессина, тем самым уменьшается реабсорбция воды и повышается ее выделение почкой.
Клеточное действие вазопрессина. Воздействие вазопрессина начинается с активации рецепторов, расположенных на базальной и латеральных мембранах клеток конечных частей дистального сегмента нефрона и собирательных трубок. Существует два типа рецепторов вазопрессина: V1и V2. Первый тип рецепторов после взаимодействия с вазопрессином активирует образование таких вторичных мессенджеров, как инозиттрифосфат и диацилглицерин. Стимуляция вазопрессином м V2-рецепторов завершается активацией фермента аденилатциклазы, в результате из АТФ образуется вторичный мессенджер — циклический АМФ (цАМФ). Это физиологически активное вещество внутри клетки вызывает ряд превращений, приводящих к увеличению проницаемости для воды плазматической мембраны, обращенной в просвет канальца. Эта мембрана называется апикальной или люминальной.
Вобычных условиях, когда в крови нет вазопрессина, люминальная мембрана непроницаема для воды, клетка не пропускает через себя воду и реабсорбция воды в собирательных трубках практически не происходит. После секреции вазопрессина в кровь
истимуляции им V2-рецепторов у внутренней поверхности базальной плазматической мембраны образуется цАМФ. Этот вторичный мессенджер должен пересечь клетку и у противоположной плазматической мембраны активировать ряд процессов, завершающихся увеличением проницаемости для воды.
Вцитоплазме клетки содержатся агрефоры — предшественники так называемых «водных каналов», которые перемещаются к люминальной мембране и встраиваются в нее. Для осуществления этого процесса необходимо участие микрофиламентов и микротрубочек. В конечном счете, цАМФ способствует встраиванию в люминальную мембрану агрегатов внутримембранных частиц; чем больше образуется цАМФ, тем большая поверхность мембраны будет занята агрегатами частиц и выше становится
проницаемость для воды. Восстановление исходной проницаемости зависит от скорости ферментативного разрушения цАМФ, эту функцию выполняет имеющаяся внутри клетки фосфодиэстераза цАМФ.
Таким образом, активация V2-рецепторов вазопрессином сопровождается образованием цАМФ и увеличением проницаемости для воды.
Выше речь шла о V1-рецепторах. Их физиологическое значение состоит в модуляции, регуляции силы ответа клетки, они обеспечивают осуществление своеобразной системы обратной связи. При регуляции водного обмена в организме ведущую роль играет лишь один гормон — вазопрессин.
Врегуляции ионного обмена обычно природа использует два гормона, оказывающие противоположное физиологическое действие, например кальцитонин, снижающий концентрацию кальция в крови, и паратгормон, увеличивающий ее.
Вслучае вазопрессина можно описать уникальный способ физиологической регуляции,
когда один гормон с помощью рецепторов двух разных типов стимулирует реакцию и
модифицирует ее. Активация V1-рецепторов и образование описанных выше вторичных мессенд-жеров способствуют снижению силы действия цАМФ. Экспериментально можно активировать только аденилатциклазу и устранить фосфоинозитидный ответ, связанный
со стимуляцией V1-рецепторов. В таком случае сила ответа на вазопрессин станет выше. Эти данные позволяют представить целостную картину регуляции водного обмена: от
возбуждения осморецепторов и секреции вазопрессина до конечных этапов внутриклеточных изменений, сопровождающихся возрастанием проницаемости мембраны для воды. Поток воды при увеличении ее реабсорбции растет в десятки раз.
Выше шла речь о процессах, происходящих в базальной мембране, цитоплазме и апикальной мембране. Такие большие физиологические изменения, сопровождающиеся возрастанием потока воды, затрагивают и внешнюю поверхность клетки. После добавления вазопрессина наблюдается экзоцитоз — выделение веществ со стороны апикальной мембраны на ее внешнюю поверхность. Среди этих веществ находятся компоненты гликокаликса, а также, вероятно, и ряд гидролитических ферментов.
А. Г. Гинецинский высказал предположение, что на одном из этапов действия АДГ в процесс вовлекается гиалуронатгидролаза, деполимеризующая гликозаминогликаны в межклеточном веществе почки. В пользу такого предположения свидетельствуют данные, что введение в кровь антисыворотки к почечной гиалуронатгидролазе уменьшает способность АДГ увеличивать проницаемость для воды и ее реабсорбцию.
12.6.8. Механизм участия почек в регуляции кислотно-основного равновесия
Почки играют важную роль в поддержании постоянства концентрации ионов водорода в крови, экскретируя кислые продукты обмена. Активная реакция мочи у человека и животных может очень резко меняться в зависимости от кислотно-основного состояния в организме. Концентрация ионов Н при крайних состояниях работы почек различается почти в 1000 раз, при подкислении мочи рН в некоторых случаях снижается до 4,5, при подщелачивании — достигает 8. Это способствует участию почек в стабилизации рН плазмы крови на уровне. 7,36. Механизм подкисления мочи основан на секреции клетками канальцев ионов водорода (рис. 12.15).
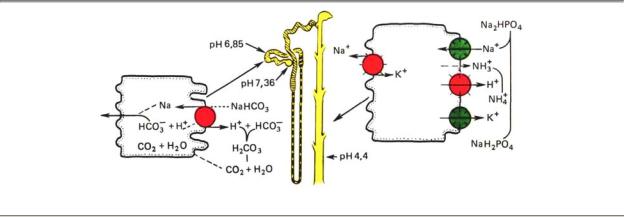
Рис. 12.15. Реакции, обеспечивающие участие почки в регуляции кислотно-основного равновесия
В апикальной плазматической мембране и цитоплазме клеток различных отделов нефрона находится фермент карбоангидраза, катализирующая реакцию гидратации оксида углерода (IV):
СО2 + Н2О ←Карбоангидраза→ Н2СО3 ← →Н+ + НСО3-
Секреция ионов Н+ создает условия для реабсорбции вместе с НСО3- эквивалентного количества ионов Na+. Наряду с Na/K ионообменным насосом и электрогенным натриевым насосом, обусловливающим перенос натрия и хлора, реабсорбция Na+ и НСО3- играет важную роль в натриевом балансе организма.
Фильтрующийся из плазмы крови бикарбонат соединяется с ионами водорода, секретированными клеткой, и в просвете канальца превращается в оксид углерода (IV) (рис. 12.15). Образование ионов водорода для секреции происходит внутри клетки, где вследствие гидратации СО2 возникшая угольная кислота диссоциирует на ионы Н+ и НСО3-. В просвете канальца ионы водорода связываются не только с бикарбонатом, но и с такими соединениями, как двузамещенный фосфат (Na2HPО4), и некоторыми другими, в результате чего увеличивается экскреция титруемых кислот (ТА-) с мочой. Это способствует выделению кислот и восстановлению резерва оснований в плазме крови.
Наконец, секретируемые ионы водорода могут связываться в просвете канальца с аммиаком, выделяющимся в клетке из глу-тамина и аминокислот и диффундирующим через мембрану в просвет канальца, в котором образуется ион аммония NH3 + Н+ → NH4+. Образование аммонийных солей нелетучих кислот способствует сбережению в организме ионов Na+ и К+, которые реабсорбируются в канальцах.
Таким образом, общая экскреция кислот почкой (UH+V) складывается из трех компонентов: количества титруемых кислот (UТАV), количества выделенного аммония (U NH4+ V) и выводимого почкой бикарбоната (U HCO3− V):
U H + V = UТАV + U NH4+ V - U HCO3− V
При питании мясом образуется больше кислот и моча становится кислой, а при потреблении растительной пищи рН мочи сдвигается в щелочную сторону. При интенсивной физической работе из мышц в кровь поступают значительные количества молочной и фосфорной кислот. В этих условиях почки увеличивают выделение кислых продуктов с мочой.
Кислотовыделительная функция почек во многом зависит от состояния кислотноосновного равновесия в организме. Так, при гиповентиляции легких происходит задержка СО2 и снижается рН крови — развивается дыхательный ацидоз, при гипервентиляции уменьшается давление СО2 в крови и растет рН крови — возникает состояние дыхательного алкалоза. Если в крови нарастает содержание ацетоуксусной и бетаоксимасляной кислот, то падает концентрация бикарбоната в крови, развивается состояние метаболического ацидоза. Рвота, сопровождающаяся потерей соляной кислоты, приводит к повышению концентрации НСО3- в крови и метаболическому алкалозу.
Легкие и почки участвуют в нормализации кислотно-основного равновесия. При дыхательном ацидозе увеличивается экскреция Н+ и реабсорбция НСО3-. Метаболический ацидоз компенсируется гипервентиляцией легких. В конечном счете почки стабилизируют концентрацию НСО3- в плазме крови на уровне 26—28 ммоль/л, а рН — на уровне 7,36.
12.6.9. Экскреторная функция почки
Почки играют ведущую роль в выделении из крови нелетучих конечных продуктов обмена и чужеродных веществ, попавших во внутреннюю среду организма. В процессе метаболизма белков и нуклеиновых кислот образуются различные конечные продукты азотистого обмена, выделяемые почкой. Обычно человек потребляет около 100 г белка в сутки. В этом количестве содержится приблизительно 16 г азота. Основными азотсодержащими веществами мочи у человека являются мочевина, мочевая кислота, креатинин, ион аммония.
Катаболизм пуриновых оснований в организме человека завершается образованием мочевой кислоты. В клетках некоторых животных имеется полный набор ферментов, обеспечивающих распад пуриновых оснований до СО2 и NH3. Мочевая кислота в почке человека фильтруется в клубочках, затем реабсорбируется в канальцах, часть мочевой кислоты секретируется клетками в просвет нефрона. Обычно у человека экскретируемая фракция мочевой кислоты довольно низка — 9,8 %, что указывает на реабсорбцию значительных ее количеств в канальцах.
Весь образующийся в течение 1 сут креатинин выделяется почками с мочой. Его источником служит креатинфосфорная кислота, от нее отщепляется фосфат, после чего в результате дегидратации креатина образуется креатинин. Его суточная продукция и экскреция зависят не только от потребления мяса с пищей, сколько от мышечной массы тела. Креатинин, как и мочевина, свободно фильтруется в почечных клубочках. Креатинин в основном экскретируется почкой, мочевина частично реабсорбируется в канальцах.
В нормальных условиях даже физиологически ценные вещества при их избытке в крови начинают экскретироваться почкой в больших количествах. Это относится как к воде и неорганическим ионам, так и органическим веществам, таким, как глюкоза, аминокислоты. В небольших количествах в мочу поступают некоторые производные продуктов гниения в кишечнике — индола, скатола, фенола, которые обезвреживаются в печени, где образуются парные соединения с серной кислотой. Это индоксилсерная, скатоксилсерная и другие кислоты.
С мочой выделяются различные биологически активные вещества и продукты их превращения, по которым в известной степени можно судить о функции некоторых желез внутренней секреции. В моче обнаружены производные гормонов коры надпочечника, эстрогены, антидиуретический гормон, катехоламины, различные витамины (аскорбиновая кислота, тиамин), ферменты (амилаза, липаза, трансаминаза) и др. В условиях патологии в моче обнаруживаются вещества, обычно в ней не выявляемые, — ацетон, желчные кислоты, белок и пр.
Помимо перечисленных существует еще очень много разнообразных веществ, непрестанно удаляемых почкой из крови. Они могут поступать в мочу и экскретироваться из организма либо после фильтрации реабсорбироваться клетками канальцев, подвергаться в них гидролизу или превращению, как, например, пептиды и измененные белки, и всасываться в кровь. О том, какие вещества выводит или разрушает почка, отчасти судят по составу крови после удаления почек. В плазме крови накапливаются не только мочевина, креатинин, мочевая кислота, но и гормоны (глюкагон, паратгормон, гастрин), ферменты (рибонуклеаза, ренин), индолы, глюкуроновая кислота, алифатические амины и многие другие вещества.
Цвет мочи зависит от величины диуреза и экскреции пигментов, он изменяется от
светло-желтого до оранжевого. Эти пигменты образуются из билирубина желчи в кишечнике, где билирубин превращается в уробилин и урохром, которые частично всасываются в кишечнике и выделяются почкой. Часть пигментов мочи представляет собой окисленные в почке продукты распада гемоглобина.
12.7. Нервная регуляция деятельности почки
Почка — один из важнейших органов, обеспечивающих гомеостаз, так как она служит исполнительным органом в цепи различных рефлексов, регулирующих постоянство состава и объема жидкостей внутренней среды. Регуляция деятельности почки, адекватная информации о состоянии внутренней среды, которая поступает в ЦНС, обеспечивается при участии эфферентных нервных волокон или эндокринних желез. Гормоны играют важнейшую роль в изменении процесса мочеобразования.
Работа почки, как и других органов, подчинена не только, безусловно-рефлекторному контролю, но и регулируется корой больших полушарий головного мозга; скорость мочеобразования может меняться условно-рефлекторным путем. В лаборатории Л. А. Орбели было показано, что прекращение мочеотделения, наступающее при болевом раздражении, может быть воспроизведено условно-рефлекторным путем, в результате стимуляции секреции АДГ нейрогипофизом. Условно-рефлекторным путем может быть вызвано и увеличение диуреза: в лаборатории К. М. Быкова было установлено, что многократное введение воды в организм собаки в сочетании с условным раздражителем также приводило к образованию условного рефлекса, при котором усиливалось мочеотделение.
Внастоящее время показано, что при участии эфферентных нервных волокон, подходящих к почке, регулируются не только ее гемодинамика и работа юкстагломерулярного аппарата, но и процессы реабсорбции и секреции неэлектролитов (глюкоза, ПАГ) и электролитов (натрий, фосфаты и др.) в канальцах. Адренэрги-ческие волокна стимулируют транспорт натрия, холинэргические волокна активируют реабсорбцию глюкозы и секрецию органических кислот. Механизм изменения мочеобразования при участии адренэргических нервов связан с активацией аденилатциклазы и образованием цАМФ в клетках канальцев. Чувствительная к катехоламинам аденилатциклаза содержится в базолатеральных мембранах клеток дистального извитого канальца и начальных отделов собирательных трубок.
Афферентные нервные волокна почки играют существенную роль в качестве информационного звена системы рено-ренальных рефлексов.
12.8.Инкреторная функция почки
Впочке образуются и выделяются в кровоток физиологически активные вещества, действующие на другие органы и ткани, а также обладающие выраженным влиянием в самой почке. К веществам, оказывающим системное и локальное действие, относятся
ренин, эритропоэтин, активная форма витамина D. В ткани почки выделяются также преимущественно локально действующие вещества, влияющие в основном на ее работу,
—простагландины и брадикинины.
Наибольшее количество данных об осуществлении инкреторной функции почки
связано с юкстагломерулярным аппаратом. Юкстагломерулярный аппарат расположен у входа в клубочек между приносящей и выносящей артериолами клубочка и частью стенки дистального канальца. В него входят гранулярные клетки афферентной артериолы, клетки плотного пятна дистального канальца и специальные клетки, контактирующие с обеими из перечисленных выше групп клеток.
Более 90 % гранул, содержащих ренин, сосредоточено в клетках стенки афферентной артериолы. Ренин является протеолитическим ферментом. Секретируемый в кровоток
ренин отщепляет в плазме от ангиотензиногена ангиотензин I. Под действием фермента от ангиотензина I, состоящего из 10 аминокислот, отщепляются две аминокислоты и образуется сильное сосудосуживающее вещество — ангиотензин II. Ангиотензин II влияет на тонус сосудов, скорость реабсорбции Na клетками канальцев, является важнейшим физиологическим стимулятором секреции альдостерона клетками коры надпочечника. Ангиотензин II очень быстро инактивируется в крови ангиотензиназами.
Описано много различных факторов, влияющих на скорость секреции ренина. Кратко охарактеризуем лишь два из них. Одним из стимулов является повышение концентрации NaCl в дистальном канальце. Это способствует секреции ренина в юкстагломерулярном аппарате данного клубочка. В нем снижается фильтрация и предотвращается возможность избыточной потери NaCl. Удержание этой соли в организме равнозначно сохранению объема внеклеточной жидкости и плазмы крови.
Другим важным стимулом служит раздражение рецепторов растяжения,
локализованных в стенке афферентной артериолы. Уменьшение ее кровенаполнения активирует выделение ренина. Гомеостатическое значение обеих реакций очевидно — вызванное секрецией ренина снижение клубочковой фильтрации приведет к сохранению циркулирующего объема и предотвратит потерю почкой избыточного количества солей натрия. Из этих данных становится понятной стратегически важная анатомическая локализация юкстагломерулярного аппарата, которая позволяет воспринимать изменения в составе канальцевой жидкости в том же самом нефроне в процессе мочеобразования и одновременно участвовать в регуляции клубочковой фильтрации и объема циркулирующей жидкости.
Таким образом, выделение ренина и образование ангиотензина имеет исключительное значение для циркуляторного гомеостаза: происходит сужение сосудов, приходит в соответствие с потребностями организма гемодинамика почки, под влиянием альдостерона усиливается реабсорбция солей натрия, способствуя сохранению объема внеклеточной жидкости в организме.
Впоследние годы были получены доказательства новой гормо-нобразующей функции почки, связанной с регуляцией обмена кальция и по своему биохимическому механизму напоминающей процессы, протекающие в коре надпочечника. Почки извлекают из крови
прогормон — 25-ОН-витамин D3, который образуется в печени, накапливают его, в митохондриях осуществляют гидроксилирование и превращение в 1,25-(ОН)2-витамин D3. Последний служит активным стероидным гормоном, стимулирующим образование Са-связывающего белка в клетках кишечника, что необходимо для всасывания Са2+. Этот же гормон способствует высвобождению Са2+ из кости.
Впочке образуется эритропоэтин, стимулирующий образование эритроцитов в костном мозгу. Особенно этот процесс активируется при гипоксии. В почке образуются кинины. В ней выделяется пептидаза — калликреин, который отщепляет кинин от кининогена во фракции альфа-глобулинов. Почечные кинины (например, брадикинин) являются сильными вазодилятаторами, участвующими в регуляции почечного кровотока
ивыделения натрия.
Вмозговом веществе почки синтезируются простагландины. Под влиянием простагландина Е2 увеличивается почечный кровоток и растет выделение Na+ без изменения клубочковой фильтрации, по-видимому, вследствие угнетения реабсорбции его
солей и воды в проксимальном канальце. Простагландин Е2 уменьшает реакцию клеток канальца на АДГ. Эти данные имеют важное значение, ибо они свидетельствуют о возможности изменения реакции на гормон под влиянием образующихся в органе физиологически активных веществ, в частности простагландинов.
Почки играют определенную роль в регуляции состояния системы свертывания крови. В них синтезируется активатор плазминогена — урокиназа. Фибринолитическая активность крови, взятой в почечной вене, значительно выше, чем в почечной артерии.
12.9. Метаболическая функция почки
Участие почки в гомеостазе белков, липидов и углеводов ранее недооценивали. Это участие не ограничено способностью к реабсорбции данных соединений или экскреции их избытка. В почке происходит образование и разрушение различных пептидных гормонов, циркулирующих в крови, образование глюкозы (глюконеогенез), превращение аминокислот, например глицина в серии, необходимый для синтеза фосфатидилсерина, который участвует в образовании и обмене плазматических мембран в различных органах.
Следует разграничить понятия метаболизм почки и метаболическая функция почки.
Метаболизм, обмен веществ в почке, обеспечивает выполнение всех ее функций, метаболическая функция почки связана с поддержанием в жидкостях внутренней среды стабильного уровня ряда углеводов, белков и липидов.
Через фильтрующую мембрану клубочка практически не проходят альбумины и глобулины, но свободно фильтруются пептиды. Тем самым в канальцы непрестанно поступают гормоны, измененные белки. Их расщепление имеет двоякое физиологическое значение — организм освобождается от физиологически активных веществ, что улучшает точность регуляции, а в кровь возвращаются аминокислоты, используемые для последующих синтезов. Имеющиеся данные указывают на возможность извлечения некоторых белков и полипептидов клетками нефрона из околоканальцевой жидкости и их последующего разрушения.
Таким образом, почка играет важную роль в расщеплении низкомолекулярных и измененных (в том числе денатурированных) белков. Это объясняет значение почки в восстановлении фонда аминокислот для клеток органов и тканей, в быстром устранении из крови физиологически активных веществ и сохранении для организма их компонентов.
Почка не только потребляет глюкозу в процессе обмена, но и обладает способностью к значительной ее продукции. В обычных условиях скорости двух последних процессов равны. Использование глюкозы для выработки энергии в почке составляет около 13 % общего потребления кислорода почкой. Глюконеогенез происходит в коре почки, а наибольшая активность гликолиза характерна для мозгового вещества почки.
Почка обладает весьма активной системой образования глюкозы, ее интенсивность на 1 г массы почки больше, чем в печени. При длительном голодании в почках образуется половина общего количества глюкозы, поступающей в кровь. Для этого используются органические кислоты, которые превращаются в глюкозу, являющуюся нейтральным веществом, что способствует одновременно регуляции рН крови. При алкалозе, напротив, снижен уровень глюконеогенеза из кислых субстратов. Зависимость скорости и характера глюконеогенеза от величины рН является особенностью углеводного обмена почки по сравнению с печенью.
Превращение различных субстратов в глюкозу, поступающую в общий кровоток и доступную для утилизации в различных органах и тканях, свидетельствует о том, что почке присуща важная функция, связанная с участием в энергетическом балансе организма.
Почка оказалась основным органом окислительного катаболизма инозитола. В ней миоинозитол окисляется в ксилулозу и затем через ряд стадий в глюкозу. В ткани почки синтезируется фосфатидилинозитол, являющийся необходимым компонентом плазматических мембран. Синтез глюкуроновой кислоты имеет большое значение для образования гликозаминогликанов, содержание которых высоко в межклеточной ткани внутреннего мозгового вещества почки и столь существенно для процесса осмотического разведения и концентрирования мочи.
Участие в обмене липидов связано с тем, что свободные жирные кислоты извлекаются почкой из крови и их окисление обеспечивает в значительной степени работу почки. Так как свободные жирные кислоты связаны в плазме с альбумином, то они не фильтруются, а их поступление в клетки нефрона происходит со стороны межклеточной жидкости. Эти