

2 курс / Нормальная физиология / Общий_курс_физиологии_человека_и_животных_Том_2_Ноздрачев_А_Д_,
.pdfтканей тела, в результате чего при одной и той же разнице температуры тела и окружающей среды величина теплоотдачи меняется. Это достигается главным образом регуляцией кровотока кожи, позицией шерсти и перьев (пилоэрекция) и испарением влаги
споверхности тела или ротовой полости.
Вморфофункциональной организации изоляции тела от температуры окружающей среды у наземных и водных гомойотермных животных существует принципиальное отличие. Это особенно заметно у арктических млекопитающих. У наземных организмов слой шерсти, выполняя основную роль термоизолятора, находится снаружи от поверхности кожи. В результате этого они не способны эффективно регулировать потерю теплоты из организма. Однако за счет меха достигается высокая термоизоляция от низких температур среды. Песец выдерживает температуру в —80°С без изменения температуры тела и видимой дрожи. При этом у арктических наземных млекопитающих, вероятно, для предупреждения перегревания во время двигательной активности существуют участки тела со слабой термоизоляцией мехом (ноги, брюшная поверхность, морда), через которые во время движения происходит эффективная теплоотдача. В покое, защищаясь от переохлаждения, животные прячут эти области, принимая характерную позу (сворачивание в клубок). Происходящее при этом уменьшение поверхности тела на 20—25% с укрытием слабо защищенных мехом областей тела является достаточно эффективным для сохранения теплоты в организме.
У водных млекопитающих термоизолирующую роль выполняет главным образом слой жира, лежащий под кожей. Предупреждение перегревания у них достигается включением кожного кровотока, обеспечивающего теплоперенос минуя термоизолирующий слой подкожного жира. Характерно, что у белого медведя, ведущего полуводный образ жизни, развит и шерстный покров, и подкожный жировой слой.
Существует одно важное обстоятельство, требующее своего разъяснения. У арктических животных имеются участки тела, практически не содержащие термоизолирующего слоя и в то же время обладающие хорошим кровоснабжением. Это ласты, плавники, подошвенная поверхность лап, ноги у птиц. Однако в указанных участках тела повышенная теплоотдача предупреждается наличием противоточных теплообменников. Устройство их таково, что притекающая артериальная кровь большую часть теплоты отдает венозной сети, которая окружает артерию, и потому притекающая с периферии венозная кровь оказывается нагретой еще до возвращения в «ядро» тела.
При угрозе перегревания у наземных гомойотермных животных на фоне увеличения кожного кровотока может включаться потоотделение. Потоотделение использует затраты тепловой энергии организма на испарение пота (2400 кДж/л воды). Эта величина свидетельствует, что путем испарения трех литров воды можно эффективно отдать всю теплоту, выработанную организмом человека в покое за 1 сут.
Некоторые животные не имеют механизма потоотделения (например, наземные хищники). Эти так называемые непотеющие животные заменяют потоотделение тепловой одышкой (полипноэ). Тепловая одышка протекает в виде сильно учащенного, но крайне поверхностного дыхания. Такой тип дыхания резко увеличивает испарение воды с поверхности верхних дыхательных путей, полости рта и языка. Следует, однако, отметить, что тепловая одышка встречается и у пойкилотермных организмов. Она обнаружена и хорошо изучена у ящериц.
Химическая терморегуляция — это реактивное повышение теплопродукции организма в ответ на действие низких температур среды. Химическая терморегуляция представляет у гомойотермных животных то средство, с помощью которого резко (в 3—5 раз) увеличивается теплообразование в организме. Увеличение теплообразования достигается за счет терморегуляционной активности мышц (сократительный термогенез) и активации специальных источников теплоты (несократительный термогенез).
Сократительный термогенез. Понятие сократительного термогенеза включает в себя терморегуляционный тонус и дрожь.
Терморегуляционный тонус является у млекопитающих и птиц аналогом позного мышечного тонуса и протекает на уровне отдельных двигательных единиц по типу низкочастотного зубчатого тетануса, близкого к режиму одиночных сокращений. Частота таких сокращений у человека, кошки, кролика и голубя находится в пределах 4—16 в 1 с. У мелких животных (массой менее 1 кг) частота этих сокращений выше и согласована с более высокой скоростью сокращения мышечных волокон. Так как сокращения отдельных двигательных единиц в мышце протекают асинхронно (т. е. не одновременно), создается внешнее впечатление тонического напряжения мышцы.
Терморегуляционный тонус возникает не во всех мышцах: вовлекаются мышцы шеи, туловища и сгибатели конечностей. Эта «топография» терморегуляционного тонуса и определяет позу, уменьшающую поверхность теплорассеивания (сворачивание в клубок).
При резком охлаждении, когда начинает падать внутренняя температура тела, включается особый процесс — холодовая дрожь. Она характеризуется периодической залповой активностью высокопороговых двигательных единиц на фоне имеющегося терморегуляционного тонуса. Низкочастотные разряды двигательных единиц во время терморегуляционного тонуса и дрожи являются крайне неэкономичными в смысле расхода энергии на каждое отдельное сокращение и тем самым дают большое высвобождение теплоты.
У крупных животных терморегуляционный тонус и дрожь являются основными источниками увеличения теплообразования в условиях холода.
Несократительный термогенез. У животных малого размера (массой тела менее 5 кг и особенно менее 1 кг) в силу высокой относительной поверхности тела при охлаждении для поддержания теплового баланса бывает недостаточно мышечной активности. Эти животные обладают другим важным источником теплообразования — так называемым
терморегуляционным несократительным термогенезом.
Важнейшим источником несократительного термогенеза является так называемая бурая жировая ткань. Наличие ее установлено не только у млекопитающих малого размера, но также у зимнеспящих животных и новорожденных, включая человека. У этих организмов отложения бурого жира обнаруживаются вокруг шеи и в межлопаточной области. Они составляют около 5 % относительной массы тела. В отличие от белого жира, где в клетке имеется одна большая жировая капля, в клетках бурого жира она разбита на мелкие капли. В бурой жировой ткани существенно больше, чем в белой, митохондрий. Бурый цвет этой ткани обусловлен большим количеством железосодержащих пигментов
—цитохромов, составляющих важную часть окислительной ферментативной системы митохондрий. Скорость окисления жирных кислот за счет такой организации в 20 раз превышает ее в белом жире, при этом идет свободное (холостое) окисление (при отсутствии синтеза и распада АТФ) с единственной целью — получения теплоты. Данный процесс осуществляется специальным полипептидом, имеющимся на внутренней поверхности мембраны митохондрий, — термогенином.
Таким образом, изменяя уровень химической и физической терморегуляции, организм может поддерживать температуру тела как интегральный показатель, отражающий соотношение теплопродукции и теплообмена с окружающей средой.
Температура тела гомойотермных животных. Сама формулировка понятия
гомойотермия предполагает сохранение устойчивой температуры только глубоко расположенных органов и тканей. В то же время поверхностные ткани претерпевают колебания температуры более существенные, чем внутренние органы. В этой связи возникло физиологически целесообразное выделение в гомойотермном организме «ядра»
—внутренних структур с устойчивой температурой (это, прежде всего, мозг и внутренние органы). «Ядро» тела окружают термоизолирующие ткани — «оболочка» (рис. 10.7).

Рис. 10.7. Температура различных областей тела человека (°С) при температуре среды 20°С (А) и 35°С (Б) Различными цветами обозначены изотермы — зоны одинаковой температуры; «ядро» тела показано наиболее темным цветом
«Ядро» и «оболочка» — чисто физиологические понятия. Например, в процессе нагревания организма, возможно, такое его тепловое состояние, когда все органы и ткани, исключая кожу, могут физиологически быть отнесены к понятию «ядро». Резкое и длительное охлаждение тела способно вызвать такое тепловое состояние, когда понятие «ядро» может распространяться лишь на мозг и внутренние органы. При этом будет наблюдаться резкое и глубокое снижение температуры тканей туловища и практически всех тканей конечностей (увеличение «оболочки» тела). Поэтому, когда речь идет о температуре тела гомойотермного организма, имеется в виду температура «ядра», т. е. внутренняя температура тела.
Наиболее строго судить о температуре «ядра» можно измеряя температуру в нижней трети пищевода — вблизи сердца. В экспериментальных физиологических исследованиях внутреннюю температуру измеряют в прямой кишке (у животных малого размера — в сигмовидной кишке), ротовой полости, тканях гипоталамуса. В медицинской практике наиболее распространенным методом является измерение температуры тела в замкнутом пространстве подмышечной впадины. Однако это требует около 10 мин (время, за которое температура подмышечной впадины приблизится к внутренней температуре тела). При этом учитывают, что измеряемая в подмышечной впадине температура обычно ниже внутренней температуры примерно на 0,5°С, так как в «ядре» тела распределение количества теплоты, а следовательно, и ее интегрального показателя — температуры — не одинаково и имеет заметное снижение к периферии. Даже в мозгу имеется поперечный градиент температур, в результате чего температура коры головного мозга может быть ниже, чем температуры ствола мозга, примерно на 1 °С. Величина этого градиента существенно зависит от размеров тела (мозга) и развития шерстного (волосяного) покрова.
Во временном отношении внутренняя температура тоже не постоянна. Температура тела большинства гомойотермных животных, ведущих дневной образ жизни, снижается к концу ночи и повышается к концу дня. У человека эта «циркадианная» периодичность выражается в колебании внутренней температуры в пределах 0,5—1°С, максимально повышаясь к 16—18 ч. и максимально снижаясь приблизительно к 4 ч утра (рис. 10.8). У животных, ведущих ночной образ жизни, наблюдается обратная динамика температуры.
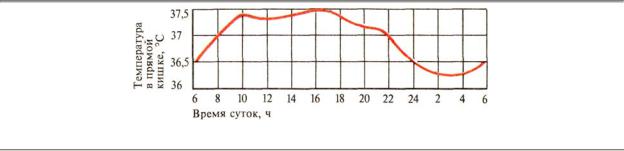
Рис. 10.8. Суточные колебания внутренней температуры тела человека
Примечательно, что именно глубокие структуры организма в покое являются главным источником теплоты, которая уже затем переносится к периферии. Это создает высокую эффективность фонового термогенеза. Так, у человека в термонейтральных условиях на долю мозга и внутренних органов, составляющих примерно 8 % относительной массы тела, приходится около 70 % всей теплопродукции организма. Естественно, что при активации химической терморегуляции начинают доминировать такие источники теплоты, как мышечная и бурая жировая ткань.
С целью поддержания нормальной температуры тела гомойотермные организмы имеют специальный механизм, управляющий химической и физической терморегуляцией, т. е. скоростью теплопродукции и теплоотдачи.
10.4.4. Терморецепция
Терморецепция появилась уже на ранних этапах филогенеза животного мира, но у пойкилотермных животных она изучена сравнительно слабо. Основные исследования свойств терморецепторов проведены на кошках, приматах и у человека. Современные представления о терморецепции сформировались только в 50—70-х гг. До настоящего времени сущность процесса терморецепции не раскрыта и единой точки зрения пока не сформулировано. Главным образом это связано с трудностями регистрации рецепторного потенциала терморецептора. Дело в том, что терморецепция осуществляется свободными окончаниями тонких сенсорных волокон типа (дельта) — С, не связанных с какимилибо специализированными образованиями. Поэтому найти такое окончание в ткани кожи, проникнуть в него микроэлектродом и зарегистрировать рецепторный потенциал в ответ на стандартные температурные стимулы пока никто из исследователей не смог. Так что основным методом изучения терморецепции остается регистрация импульсов, отводимых от афферентных волокон кожного нерва.
Исследования показали, что терморецепторы представлены в «оболочке» тела: коже, подкожных тканях (в том числе кожных и подкожных сосудах) — это периферические рецепторы, — и в ЦНС: наибольшая плотность их обнаружена в медиальной преоптической области гипоталамуса — это центральные нейроны (термосенсоры).
У высших млекопитающих, в том числе и у человека, установлено существование специфических терморецепторов, крайне мало чувствительных к нетемпературным стимулам. Эти терморецепторы подразделяются на две популяции: холодовые и тепловые рецепторы. Характерным для обеих популяций является то, что при постоянной температуре кожи они разряжаются тонически с частотой, зависящей от температуры (так называемая статическая активность терморецепторов).
Средние частоты разрядов для обеих популяций обладают своими максимумами (рис. 10.9). Для холодовых терморецепторов максимальная частота разрядов обнаружена в пределах 20—33°С (средняя 26°С), для тепловых — в пределах 40—46°С (средняя 43°С). При медленном изменении температур по обе стороны температурных максимумов активность холодовых и тепловых терморецепторов снижается (так называемый
колоколообразный характер статической активности терморецепторов).

Рис. 10.9. Активность холодовых (1) и тепловых (2) терморецепторов обезьян при постоянной температуре кожи
Следует заметить, что ощущение горячего (жжение), возникающее при температуре свыше 45°С, воспринимается другими рецепторами — горячевыми (или рецепторами жжения). Эти рецепторы относятся к категории полимодальных ноцицепторов и являются как бы промежуточной популяцией между терморецепторами и ноцицепторами.
Температурные диапазоны активности холодовых и тепловых рецепторов широко перекрываются, поэтому их нельзя отличить по статической активности. Отличие устанавливают по реакции на резкое изменение температурного воздействия. Холодовые рецепторы реагируют резким кратковременным учащением разрядов на охлаждение и кратковременным прекращением их — на нагревание. Тепловые рецепторы, наоборот, повышают активность на резкое нагревание.
Установлено, что среди периферических терморецепторов количественно преобладают холодовые терморецепторы, тогда как подавляющее большинство терморецепторных нейронов гипоталамуса отвечает учащением разряда на нагревание, т. е. относится к тепловым сенсорам.
10.4.5. Центральный механизм терморегуляции
Общепринято, что основной центральный механизм терморегуляции (так называемый центр терморегуляции) локализован в структурах гипоталамуса.
Еще сравнительно недавно выделяли центр теплоотдачи, локализованный в переднем гипоталамусе, и центр теплопродукции, локализованный в заднем гипоталамусе, которые гипотетически находятся в реципрокных отношениях. Возбуждение первого приводит к усилению теплоотдачи и уменьшению теплопродукции (падение температуры тела), возбуждение второго — к усилению теплопродукции и уменьшению теплоотдачи (повышение температуры тела). Однако дальнейшие исследования не подтвердили наличие рецеп-рокности указанных структур гипоталамуса.
По современным представлениям, терморегуляция осуществляется распределенной системой, основной частью которой является гипоталамический терморегуляционный механизм. Сигнализация от периферических терморецепторов, переключаясь в структурах задних рогов спинного мозга, адресуется к сегментарным соматическим и автономным механизмам своего спинального уровня, а также поступает по восходящим путям спинного мозга в головной мозг. Главными проводниками температурной чувствительности в головной мозг являются спиноталамический и спиноретикулярный тракты. Причем спиноталамический тракт состоит из компонентов разного филогенетического возраста — неоспиноталамического и па-леоспиноталамического трактов.
Неоспиноталамический тракт относится к категории лемнисковых систем и обеспечивает контралатеральную соматотопическую проекцию (локализацию) рецепций температуры (как, впрочем, и боли) через таламические переключающие ядра в кору головного мозга. Палеоспиноталамический и спиноретикулярный тракты относятся к
категории экстралемнисковых путей. Температурная информация по этим древним путям диффузно через ретикулярную формацию и неспецифические ядра таламуса поступает не только в ассоциативные зоны коры головного мозга, но и в структуры гипоталамуса. Сигнализация от периферических терморецепторов адресуется в передний гипоталамус (медиальную преоптическую область), где происходит сравнение этих сигналов с уровнем активности центральных термосенсоров (отражающих температурное состояние мозга). Эта интеграция сигналов, характеризующих центральную и периферическую температуру тела, и обеспечивает выработку структурами заднего гипоталамуса сигналов, управляющих химической и физической терморегуляцией.
В комфортных (термонейтральных) условиях тепловой баланс, обеспечивающий поддержание температуры тела на нормальном уровне, не нуждается в коррекции специальными механизмами терморегуляции. Для разных видов комфортная температура среды различна. Однако даже для отдельного индивида она зависит не только от самой температуры, но меняется в зависимости от степени влажности воздуха, конвективных потоков и теплопроводности среды. Так, газовая среда, где азот заменен гелием, примерно в 7 раз теплопроводнее, чем воздушная, а водная среда приблизительно в 20 раз теплопроводнее, чем воздушная. В проточной воде (из-за высокой конвективности) охлаждающее или перегревающее действие на организм в 50—100 раз больше, чем на воздухе.
Температура среды ниже комфортной вызывает увеличение активности холодовых периферических терморецепторов. Эта «холодовая» информация повышает тонус эфферентных структур заднего гипоталамуса, в результате чего через активацию симпатической нервной системы повышается тонус кожных и подкожных сосудов (рис. 10.10). Уменьшение кровотока в «оболочке» тела, связанное с повышением тонуса указанных сосудов, приводит к повышению термоизоляции организма и сохранению теплоты за счет уменьшения теплоотдачи. Вместе с этим возникшее в связи с охлаждением усиление симпатического тонуса вызывает пиломоторный рефлекс — симпатическое сокращение гладкомышечных пучков, поднимающих шерстный (волосяной, перьевой) покров, в результате чего термоизолирующая роль шерстного (перьевого) покрова у животных возрастает.

Рис. 10.10. Механизмы теплоотдачи организма в условиях холода (А) и тепла (Б): 1 —
перераспределение крови между сосудами внутренних органов (I) и сосудами поверхности кожи (2); II — перераспределение крови в сосудах кожи; 3 — кожа; 4 — кровеносные сосуды; 5 — артериовенозные анастомозы; красными стрелками обозначено движение крови, синими — пути теплоотдачи
Параллельно возникновению реакции теплоконсервации эфферентные структуры заднего гипоталамуса активируют систему регуляции позного мышечного тонуса (появление терморегуляционного тонуса и дрожи), что приводит к увеличению выработки теплоты в организме (сократительный термогенез).
Высвобождение норадреналина и адреналина симпатической нервной системой и надпочечниками через тканевые бета-адрено-рецепторы стимулирует энергообмен во всех тканях, в том числе и в бурой жировой ткани у тех организмов, которые ее имеют (несократительный термогенез). Эффективность адренэргической стимуляции теплообразования потенциируется под действием тиреоидных гормонов (трииодтиронина и тироксина), выделение которых при охлаждении также усиливается.
Согревание животного уменьшает активность холодовых периферических терморецепторов, вызывая уменьшение тонуса эфферентных структур гипоталамуса. Снижение эфферентных влияний центра терморегуляции приводит к уменьшению симпатических влияний на кожные и подкожные сосуды, что сопровождается увеличением кожного кровотока вследствие «пассивного» сосудорасширения, уменьшается адренэргическая и тиреоидная активация энергообмена (рис. 10.10). Снижение эфферентных влияний центра терморегуляции вызывает и уменьшение мышечного тонуса, и термогенеза, связанного с ним.
В условиях перегревания организма ведущая роль в возникновении реакций терморегуляции принадлежит возрастающей активности тепловых нейроновтермосенсоров медиальной преоптической области. Активность этих тепловых нейронов включает две реакции.
Во-первых, активируются особые структуры симпатической нервной системы, управляющие потоотделением через холинэргические нервные волокна, в результате чего резко увеличивается теплоотдача испарением. У непотеющих животных эту реакцию заменяет высокочастотная активация диафрагмы, приводящая к тепловой одышке.

Во-вторых, резко тормозится активность всех мышц, кроме диафрагмы, включая и активность межреберных мышц. Суммарная активность диафрагмы во время тепловой одышки не увеличивается. Залпы инспирации становятся чаще, но пропорционально менее длительны. Лишь изредка они прерываются нормальным дыхательным циклом для полной вентиляции легких. В условиях сильного перегревания двигательная активность млекопитающего подавлена полностью (поза распластывания); человек при этом не способен совершать произвольные двигательные акты, вызываемые волевым усилием. Это подавление двигательной активности направлено на минимизацию мышечного термогенеза при угрозе перегревания.
Резкое подавление функциональной активности у животных при перегревании связано с тем, что летальная температура для организма в этой ситуации в среднем лишь на 6°С превышает нормальную (табл. 10.4). Границы низких летальных температур менее определенны из-за большей устойчивости гомойотермных организмов к переохлаждению в связи со снижением энергообмена тканей. Явление уменьшения температуры ниже нормальной более чем на 2°С называют гипотермией. У человека падение температуры тела ниже 35°С приводит к нарушению поведения. При температуре тела около 31°С наступает потеря сознания, а при температуре около 24—26°С — смерть из-за нарушения автоматии сердца. Другие млекопитающие более устойчивы к переохлаждению организма: собаки погибают при снижении внутренней температуры до 18—20°С, кошки
— до 14—16, крысы — до 13—15°С.
Таблица 10.4. Приближенные значения нормальной и летальной внутренней температуры у млекопитающих и птиц (по К. Шмидт-Ниельсону, 1982)
Группа животных |
Нормальная |
Летальная |
|
температура |
температура тела, °С |
|
тела, °С |
37 |
Однопроходные (ехидна) |
30—31 |
|
Сумчатые |
35—36 |
40—41 |
Насекомоядные (еж) |
34—36 |
41 |
Человек |
37 |
43 |
Высшие млекопитающие |
36—38 |
42—44 |
Птицы (кроме воробьиных) |
39—40 |
46 |
Птицы (воробьиные) |
40—41 |
47 |
10.4.6. Температурная адаптация
Продолжительное (около месяца) пребывание гомойотермного животного в перегревающих или в переохлаждающих условиях микроклимата приводит к повышению эффективности механизмов защиты от перегревания и соответственно от переохлаждения. Это явление называют температурной адаптацией (более точное название —
температурная акклимация).
Тепловая адаптация сводится главным образом к повышению эффективности механизма потоотделения, что достигается за счет повышения чувства жажды при незначительных потерях воды и снижения порога потоотделения на перегревание.
Холодовая адаптация заключается в увеличении теплоизолирующих свойств меха и накопления подкожного жира, а также в фоновом повышении тканевого энергообмена (термогенеза) за счет увеличения количества тканевых бета-адренорецепторов. В результате увеличения количества бета-адренорецепторов в мышцах и их активации норадреналином сократительный акт становится менее экономичным, а потому терморегуляционный тонус и дрожь у адаптированных к холоду животных дают больший выход теплоты. Вероятно, эти же механизмы лежат в основе сезонных перестроек системы терморегуляции.
Поддержание высокого уровня метаболизма тела в условиях холода — дорогостоящий механизм, особенно для мелких животных, у которых фоновый энергообмен напряжен и
связан с большим потреблением пищи. Поэтому у ряда мелких гомойотермных организмов эволюционно сформировался особый механизм защиты от холода — зимняя спячка (гибернация). Зимняя спячка — это переход на низкий уровень энергообмена путем снижения температуры тела примерно до + 5°С. Зимней спячке подвержены грызуны (хомяки, суслики, сурки, сони), насекомоядные (ежи), рукокрылые (летучие мыши) и др. Перед переходом к зимней спячке гибернирующие животные запасают подкожный жир, энергия химических связей которого и тратится на минимальный подогрев организма во время спячки.
Бурые медведи не являются истинно гибернирующими животными. Они впадают в состояние зимнего сна, лишь слегка понижая температуру тела (примерно до 33—31°С) и способны быстро выходить из состояния сна.
10.5. Питание
Питание — это процесс усвоения организмом веществ, необходимых для построения и обновления тканей его тела, а также для покрытия энергетических затрат.
В целом эволюция пищевых потребностей животных организмов включала в себя процесс ограничения собственного синтеза ряда соединений с одновременным расширением потребления органических соединений определенных типов. Это привело к выделению целой группы веществ, незаменимых для высших животных и человека, т. е. необходимых для обмена веществ, но не синтезируемых самостоятельно.
Использование пищевых средств, состоящих в основном из сложных соединений растительного или животного происхождения, для энергетических или пластических нужд организма возможно только после гидролиза этих средств и превращения в сравнительно простые соединения, лишенные видовой специфичности. Пищевые потребности разных видов животных различны в зависимости от того, какие пищевые вещества организм способен синтезировать и какие должны поступать извне. И все же в основном различия в пищевых потребностях обусловлены способами переваривания (гидролиза) пищи. Это связано с тем, что у высших животных организмов промежуточные процессы обмена веществ протекают сходным образом.
Вобмене веществ (метаболизм) и энергии различают два направленных процесса: анаболизм и катаболизм. Под анаболизмом понимают совокупность процессов, направленных на построение структур организма главным образом через синтез сложных органических веществ; под катаболизмом — совокупность процессов распада сложных органических соединений и использование образовавшихся при этом сравнительно простых веществ в процессах энергообмена. В основе анаболизма и катаболизма лежат соответственно процессы ассимиляции и диссимиляции, которые в организме взаимосвязаны и в нормальном организме сбалансированы (см. рис. 10.1).
Вцелом потребности животных достаточно однородны: они нуждаются в сходных по структуре питательных веществах для энергообмена; в веществах типа аминокислот, пуринов и некоторых липидов для построения сложных белковых молекул и клеточных структур; в специальных катализаторах обмена веществ и стабилизаторах клеточных мембран; в неорганических ионах и соединениях для физико-химических процессов в организме и, наконец, в универсальном биологическом растворителе — воде — для создания среды клеточного обмена веществ.
Вконечном итоге в состав пищи высокоорганизованных организмов входят органические вещества, подавляющая часть которых относится к белкам, липидам и углеводам. Продукты их гидролиза — аминокислоты, жирные кислоты, глицерин и моносахара — тратятся на энергообеспечение организма. В процессах энергообмена аминокислоты, жирные кислоты и моносахара взаимосвязаны общими путями их превращения. Поэтому как энергоносители пищевые вещества могут взаимозаменяться в соответствии с энергетической ценностью (правило изодинамии).
Энергетическую (калорическую) ценность пищи оценивают по количеству тепловой энергии, высвобождающейся при сгорании 1 г пищевого вещества (физиологическая теплота сгорания), которую выражают традиционно в килокалориях или по СИ — в джоулях (1 ккал = 4,187 кДж). Расчеты показали, что энергетическая ценность жиров (38,9 кДж/г; 9,3 ккал/г) в два раза выше, чем белков и углеводов (17,2 кДж/г; 4,1 ккал/г). Белки и углеводы имеют одинаковую энергетическую ценность и могут заменяться 1:1 в весовом соотношении.
Для поддержания стационарного состояния организма общие затраты энергии должны покрываться поступлением пищевых веществ, несущих в своих химических связях эквивалентный запас энергии. Если количества поступающей пищи для покрытия энерготрат недостаточно, то энерготраты компенсируются за счет внутренних резервов, главным образом — жира. Если же масса поступающей пищи по энергоносителям превышает расход энергии, то идет процесс запасания жира независимо от состава пищи.
Однако следует всегда помнить, что эти три источника энергии являются и
пластическим материалом животного организма. Поэтому длительное исключение одного из трех питательных веществ из пищевого рациона и замена энергетически эквивалентным количеством другого вещества недопустимы.
10.5.1. Рациональное питание
Под рациональным питанием понимают достаточное в количественном и полноценное в качественном отношении питание. Основа рационального питания — сбалансированность, т. е. оптимальное соотношение компонентов пищи (аминокислот, полиненасыщенных жирных кислот, фосфатидов, стеринов, жиров, Сахаров, витаминов, минеральных солей, органических кислот и т. д.). Насчитывается около 60 пищевых веществ, требующих сбалансированности. Рациональное питание обеспечивает оптимальное поступление пластических, энергетических и регуляторных веществ, необходимых для нормальной жизнедеятельности организма. Однообразное питание, приводящее к исключению отдельных компонентов сбалансированного пищевого рациона, вызывает нарушение обмена веществ.
Для человека и многих плацентарных животных сбалансированное питание должно включать белки, жиры и углеводы в весовой пропорции примерно 1:1:4. Это дает нормирование суточной калорийности пищевого рациона за счет белков: 15% суточной калорийности (причем белки животного происхождения должны составлять не менее половины). Доля жиров при указанном соотношении составляет примерно 30 % суточной калорийности (причем 75—80% жира должны составлять животные жиры). Энергетическая доля углеводов при таком соотношении должна составить 55%.
10.5.2. Белки пищи
Особое значение в сбалансированном питании имеют белки. Белки, как известно, состоят из аминокислот. Аминокислоты являются как источником синтеза структурных белков, ферментов, гормонов белковой и пептидной природы и т. д., так и источником энергии.
Характеристика белков, входящих в состав пищевого рациона, включает не только энергетическую ценность, но и спектр аминокислот. Это важный показатель, так как даже после завершения роста организм постоянно обновляет белковые структуры. Средний полупериод распада белков тела — условная величина и резко отличается у разных видов. Так, полупериод распада белка тела человека составляет 80 сут, а тела крысы — всего лишь 17 сут. При этом следует иметь в виду, что многие белковые структуры даже у одного вида обновляются с разной скоростью. Наиболее медленно обновляются мышечные белки. Полупериод распада их у человека составляет примерно 180 сут, а у